
去年のサイエンス誌の報告ですが、車のゴムタイヤに添加される酸化防止剤6PPDが、オゾンと反応してキノン化(ベンゼン環に酸素が2個つくこと)されて道路に散乱し、降雨で河口に流されて、銀鮭が大量死することが分かったそうです。科学者たちの執念の成果です。

GIGAZINE.NET
科学者たちが「雨が降ると川のギンザケが大量死する」という謎を20年かけて解き明かす
by Bureau of Land Management Oregon and Washington


NHKスペシャル・2030 未来への分岐点(1)「暴走する温暖化脱炭素への挑戦」に続いて、2021年2月7日(日)に2030 未来への分岐点 (2)「飽食の悪夢〜水・食料クライシス〜」が放送されました。先進国の飽食が、世界中に「飢餓のパンデミック」を拡大させるという話です。番組では俳優の森七菜さんが2050年の日本で食糧危機に直面する女性を演じました。日本で一年間に出される食品廃棄物を世界に分配すれば、飢餓問題の多くを解決すると言われています。現在の食料システムを2030年までに改善できなければ、暴動が発生し破滅を回避できなくなると研究者たちは指摘しています。
米国メリ-ランド大学の理論環境学者のSafa Mote博士は、2014年に「人間と自然の動力学(=Human And Nature Dynamics)~社会の崩壊か持続における不平等と資源利用のモデル化~」と題して、論文を発表しました(Ecological Economics 101(2014)90-102)。このモデルをHANDYモデルと呼びます。
Safa Mote博士は、自然から収奪する富の蓄積と富の再分配をモデル化して、平等社会と不平等社会における人口変動を予測しました。Safa Mote博士はHANDYモデルにおいて
ことを示しました。ここでは、HANDYモデルの詳細について解説します。

Safa Mote博士が2014年に提案した「人間と自然の動力学(Handyモデル」」は、一般の人口Xc、エリ-トの人口XE、自然資源量y、富の総量wの4つの量の時間発展を4つの連立微分方程式で表したモデルです。その名の通り、このモデルは文明存続を議論するための最もお手軽なモデルになっています。
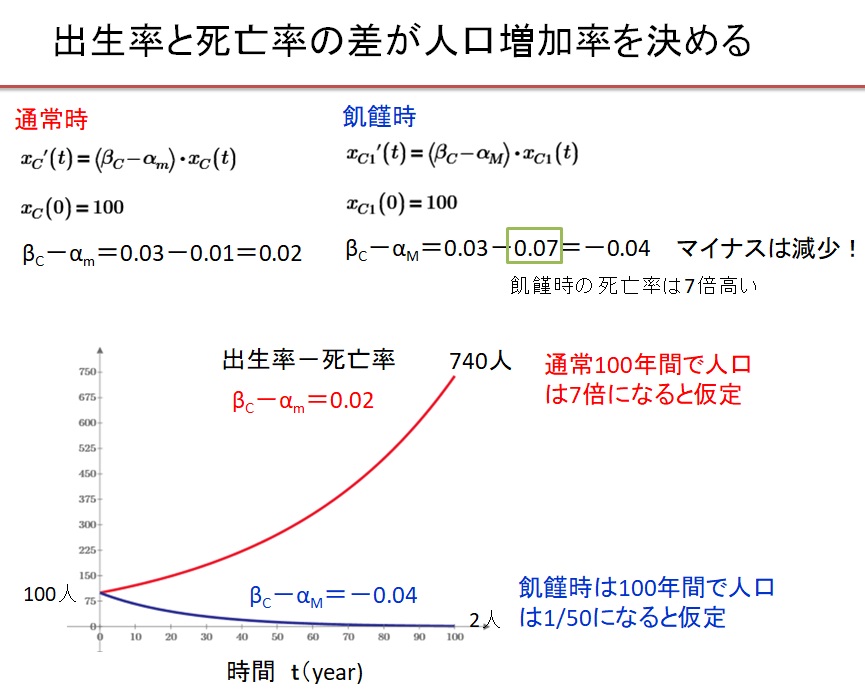
(1)一般人の人口Xc
人口増加は、出生率をβ、死亡率をαとすると
と表されます。ここで
αc=αc(Xc、XE、w)、β=出生率定数
です。α、βがともに定数の場合、一般人の人口変動は
と書き表されます。出生率が死亡率より大きい場合(β-α>0)、一般の人口Xcは指数関数的に増加します。逆に出生率が死亡率より小さい場合(β-α<0)、一般の人口Xcは指数関数的に減少します。
(2)エリ-トの人口XE
エリ-トの人口も同様に微分方程式
αE=αE(Xc、XE、w)、β=出生率定数
と表されると仮定します。
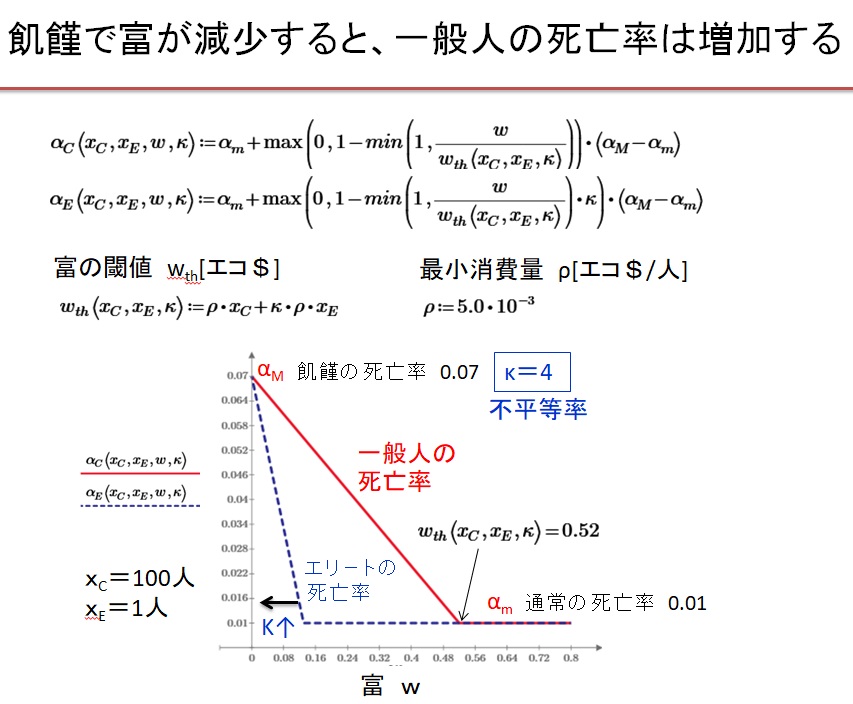
一般に死亡率をαは、人口や富の量に依存します。富の量が人口を養うのに十分であれば、一定になりますが、富の量が閾値wthより小さくなると、死亡率は増大します。一般人とエリ-トでは死亡率αの富の総量wに対する依存性が異なります。
(3)富の閾値wthと不平等率kと死亡率αの富w依存性
ρ[$/人]を一般人一人当たりの最小消費量とすると、富の閾値は
・wth(Xc、XE、k)=ρ・Xc+k・ρ・XE
と表されると仮定します。ここで因子kは不平等率です。エリ-トの最小消費量は一般人のk倍と仮定されています。kの値によって社会を3つに分類できます。
1)平等社会 Egalitarian society k=0、エリ-トなし
2)階級社会 Equitable society k=1、不労所得階級あり
3)不平等社会 Unequal society k>1、エリ-トあり
具体的に、このモデルでは通常の死亡率はαm=0.01、飢饉時の死亡率はαM=0.07としています。死亡率αは、富が閾値を下回ると0.01から0.07に富に比例して増大すると仮定します。一般人とエリ-トでは富の閾値が異なります。エリ-トの富の閾値は小さいので、エリ-トの死亡率は殆ど最小値0.01に固定されています。つまり富の総量wが閾値より減少すると、一般人の死亡率は減少し始めますが、エリ-トの死亡率は低いまま保たれます。
(4)資源量y
限られた食物環境にある生物の増殖を議論するのには、ロジスティック方程式が用いられます。資源量yは、ロジスティック方程式
・dy/dt=γy(λ-y)-δ・Xc・y
に従うと仮定します。ここでλは人間の収奪がないときの最大資源量です。λの単位は$(エコドル)です。γ[1/t$]は単位時間の自然の再生率です。γ=0.01のとき10年で再生します。δ[1/t人]は一人当たりの人間が1年間に自然から収奪する収奪率です。収奪率δがゼロだと富が蓄積されません。仮にy(0)=λ/1000とすると、δ=0のときは、資源量y(t)は、S字型の再生曲線を描いて増加し、最大資源量λで飽和します。なぜならλ≫yのときは、指数関数的に増大し、yが増大してλに接近するとyは一定値λになるからです。
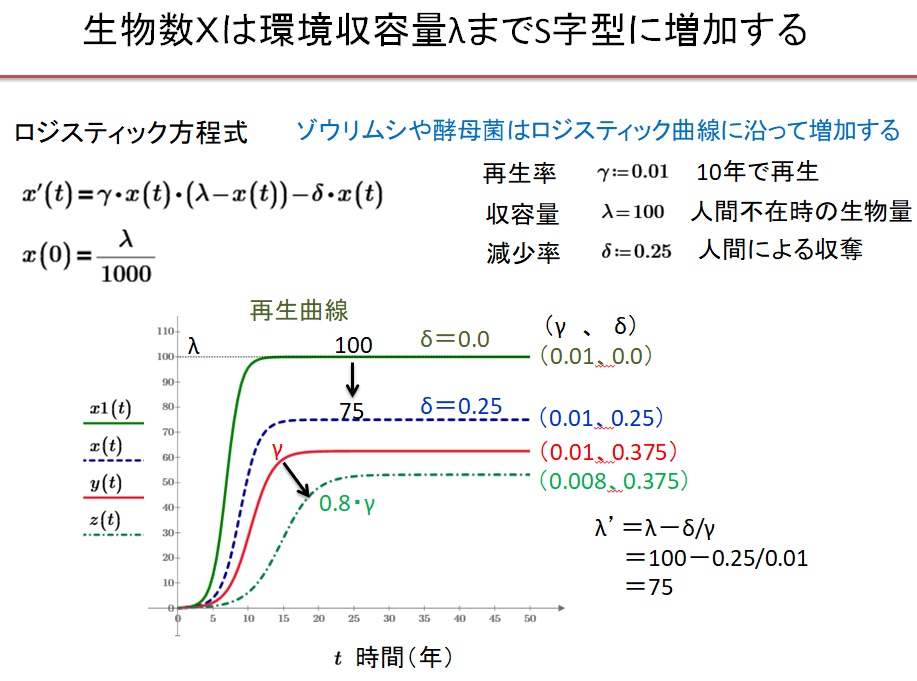
δがゼロでないときは
・dy/dt=γy(λ-δXc/γ-y)=γy(λ’-y)
と書けます。資源量y(t)は、S字型の再生曲線を描いて増加しλ’で飽和します。
λ=100、γ=0.01、δ=0.0025、Xc=100のときは
・λ’=λ-δXc/γ=100-0.0025*100/0.01=100-25=75 <100=λ
となります。
y<<1の時は、yの2次の項を無視して
・dy/dt=(γλ-δXc)y
と近似できます。収奪率δがγλ/Xcより大きくなると、資源量yは減少し、人類は滅んでしまうことが分かります。収奪率δが
・δ[1/t人]=γ[1/t$]・λ[$]/Xc[人]=0.01・100/100=0.01
のとき、資源量yは一定になります。
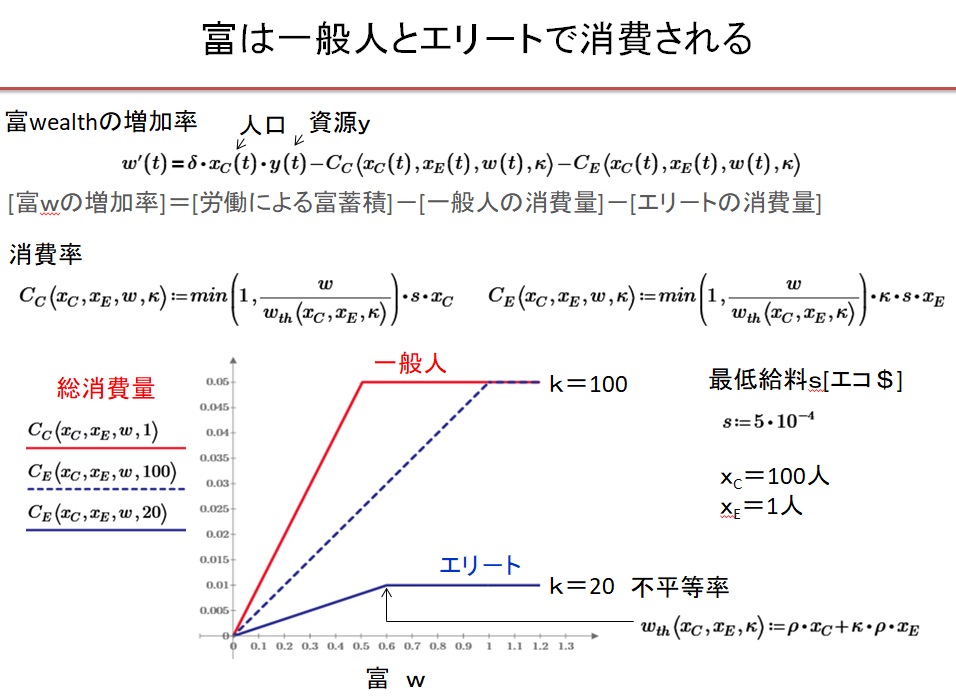
(5)富の総量w
富の総量wは、
・dw/dt=δXc・y-Cc(Xc、XE、w、k)-CE(Xc、XE、w、k)
に従うと仮定します。1年間の富の増加量は、自然から得た収奪量から一般人による富の消費量Cc[$/t]とエリ-トによる富の消費量CE[$/t]を引いた値になります。自然からの収奪量と富の消費量が一致する循環社会では、富は一定の値に保たれ、人口も安定します。
最低給料をs[$/人]とすると、一般人とエリ-トの富の消費量は
・Cc(Xc、XE、w、k)=min(1,w/wth)・s・Xc
・CE(Xc、XE、w、k)=min(1,w/wth)・s・k・XE
と表されると仮定します。エリ-トの消費量の場合は不平等率kがかかります。
ここで富の閾値は
・wth(Xc、XE、k)=ρ・Xc+k・ρ・XE
でした。w>wthの平時では、Ccは最低消費量
・Cc(Xc、XE、w、k)=s・Xc
となり、Ccはwに依存しません。w<wthの飢饉の時は、
・Cc(Xc、XE、w、k)=w/wth・s・Xc
となり、Ccはwに比例します。富の総量wがwthより小さくなる飢餓状況では、人の消費量はwが減るにつれて減少することになります。不平等率kが大きいほどエリ-トの消費量は大きくなります。
(6)初期状態
簡単のため、初期状態は
・一般人の人口:Xc(0)=100[人]、
・エリ-トの人口:XE(0)=1[人]
・資源量:y(0)=100[$]、
・富の総量:w(0)=0[$]
と仮定しています。
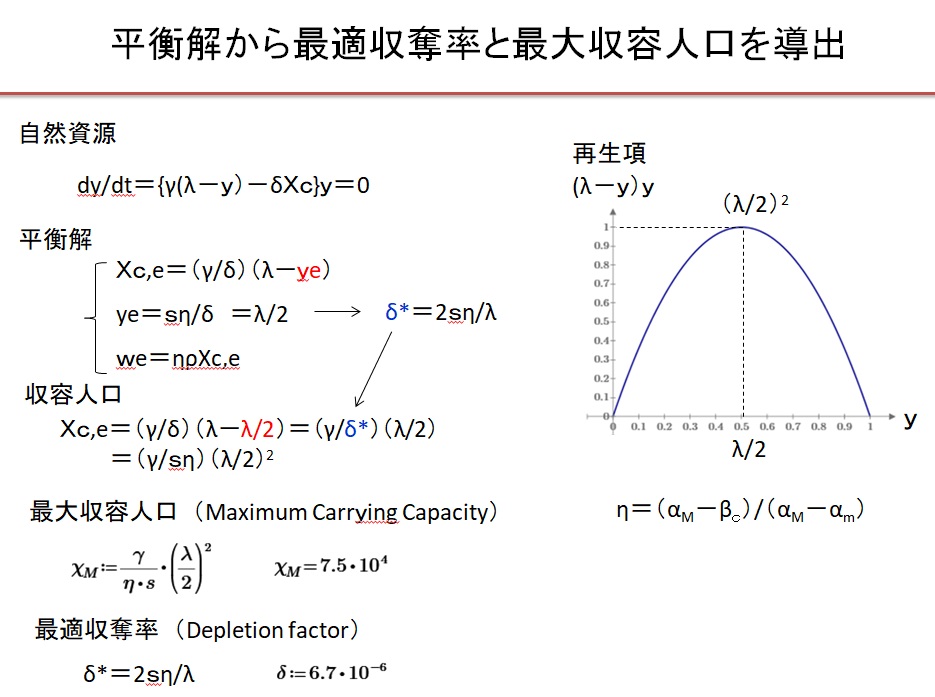
2.平衡状態の人口、資源、富の量 XE=0の場合
ここでは、簡単のためエリ-トがいない平等社会での
・dXc/dt=0,dy/dt=0、dw/dt=0
なる平衡状態(定常状態)の解Xce、ye、weを考えます。以下に平衡解の導出方法を示します。

パラメ-タηを
・η=(αM-βc)/(αM-αm)
と定義すると、結局、平衡時の資源量yeは
・ye=sη/δ (=λ/2)
と書けます。平衡時の人口Xce、富weは
・Xce=γ/δ・(λ-ye)
・we=ηρXce
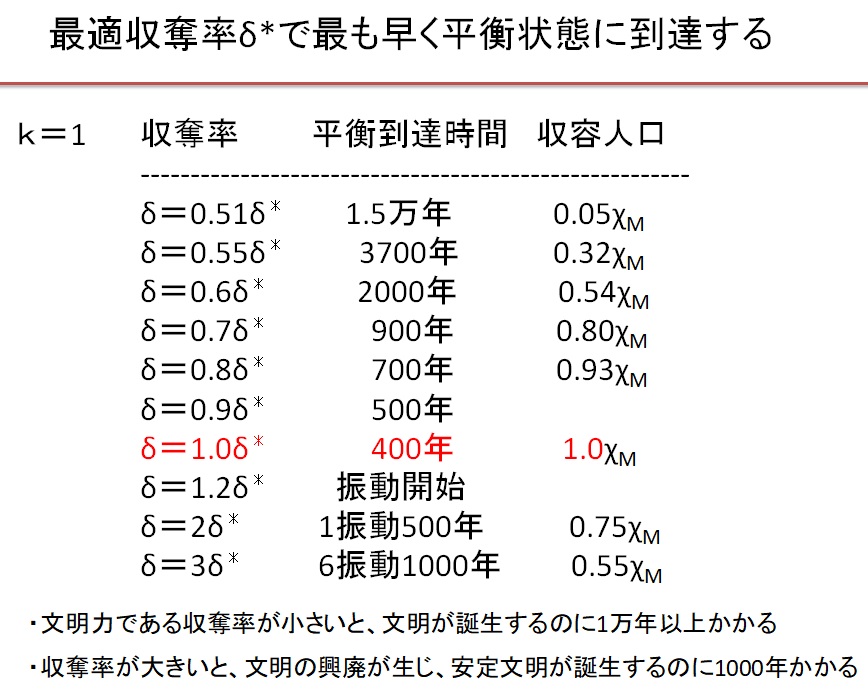
と書けます。平衡時の資源量ye=λ/2のとき、再生項y(λ-y)が最大値λ2/4になるので、このときの収奪率を最適収奪率(Optimal Depletion Ratio)
・δ*=2sη/λ=6.7×10-6
と呼びます。δ=δ*のとき、最大収容量(Maximum Carrying Capacity)
・XM=γ/δ*・(λ-λ/2)=γ/ sη(λ/2)2=7.5×104
が得られます。
3.計算に用いたパラメ-タ

4.Equitable社会の持続可能性の収奪率依存性について
少数の不労者はいるが、不労者の消費量は一般人と同じ(k=1)である階級社会をEquitable societyと言います。kは不平等率です。
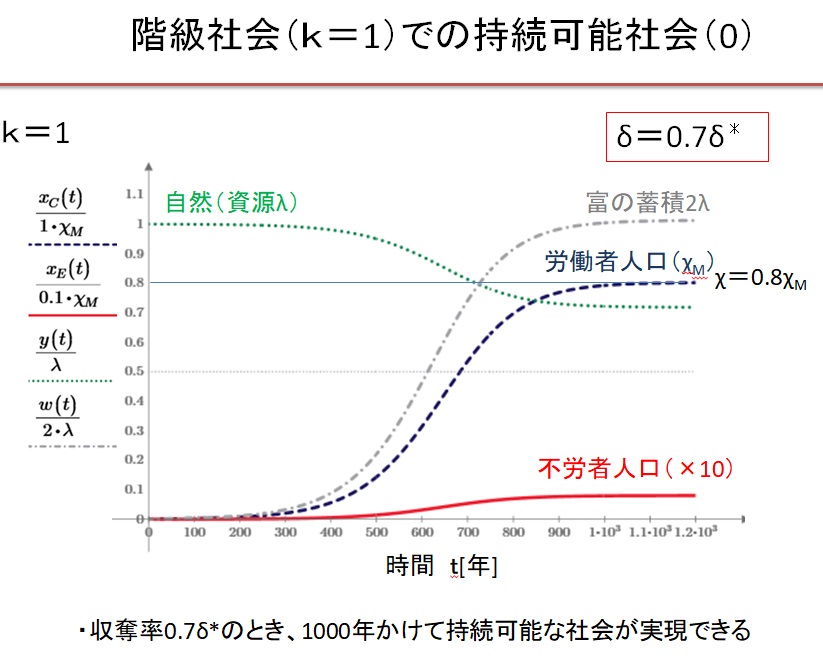
(1)k=1の階級社会 δ=0.7・δ*の場合
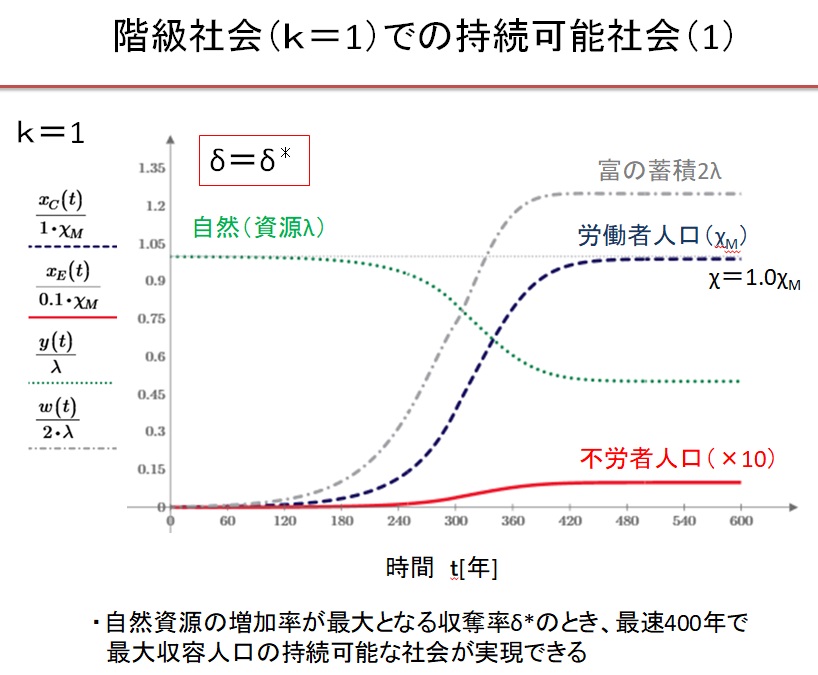
(2)k=1の階級社会 δ=1.0・δ*の場合
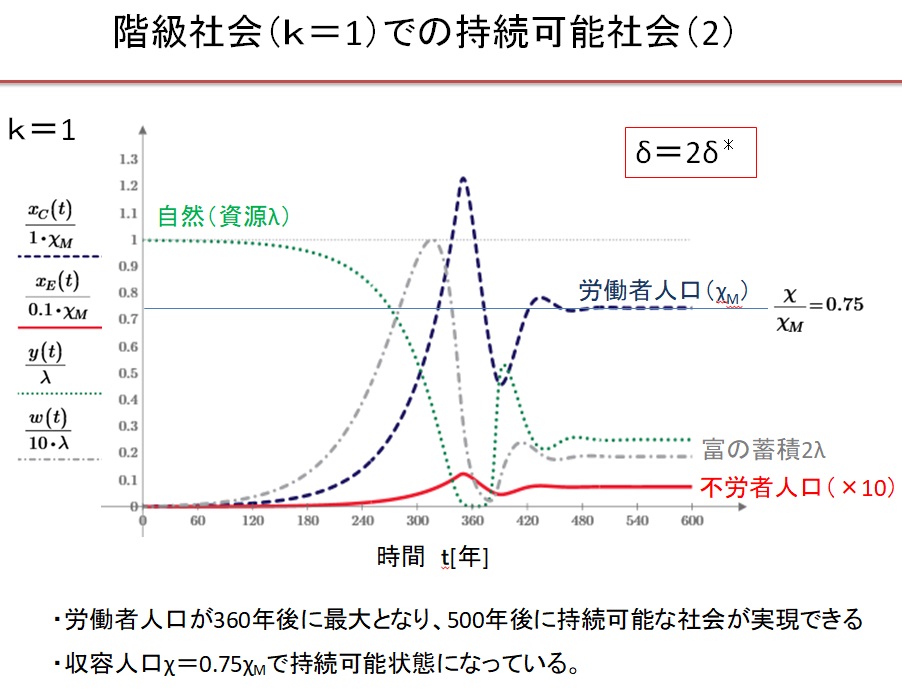
(3)k=1の階級社会 δ=2.0・δ*の場合
(4)k=1の階級社会 δ=3.0・δ*の場合
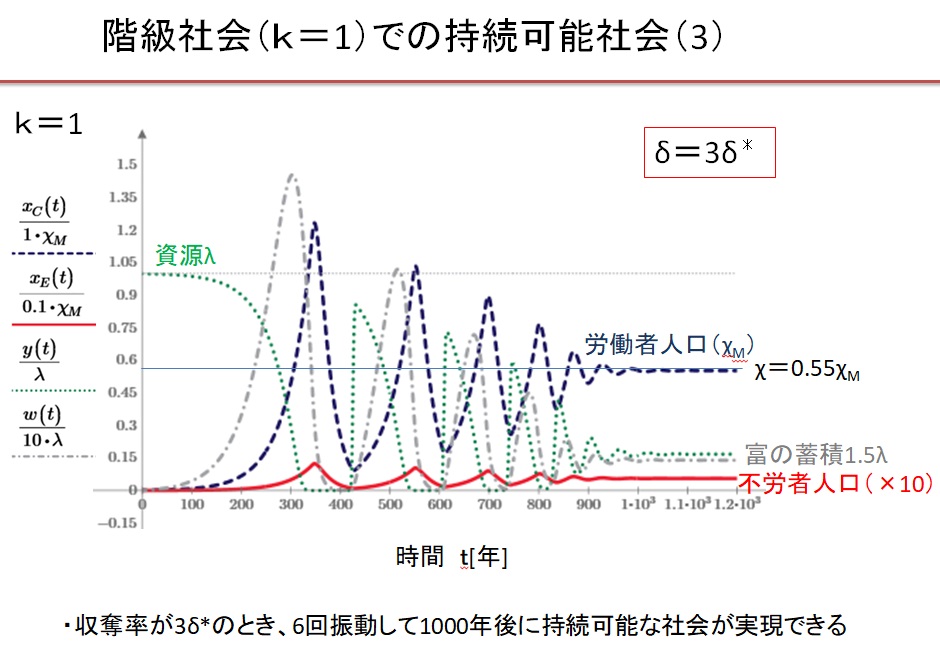
(5)Equitable 社会のまとめ
Equitable 社会では、0.55δ*~3δ*の広い収奪率で持続可能な文明が実現します。収奪率が最適収奪率の0.55倍の場合は、人口増加が遅く、収容人口は最大値の1/3になります。最適収奪率δ*のとき、最速400年で持続可能な社会が実現し、収容人口は最大になります。収奪率が最適収奪率の2倍になると、収容人口は最大値の3/4に減少します。収奪率が最適収奪率の2倍以上になると、振動現象が現れ、持続可能な文明に到達するのに1000年以上を要します。
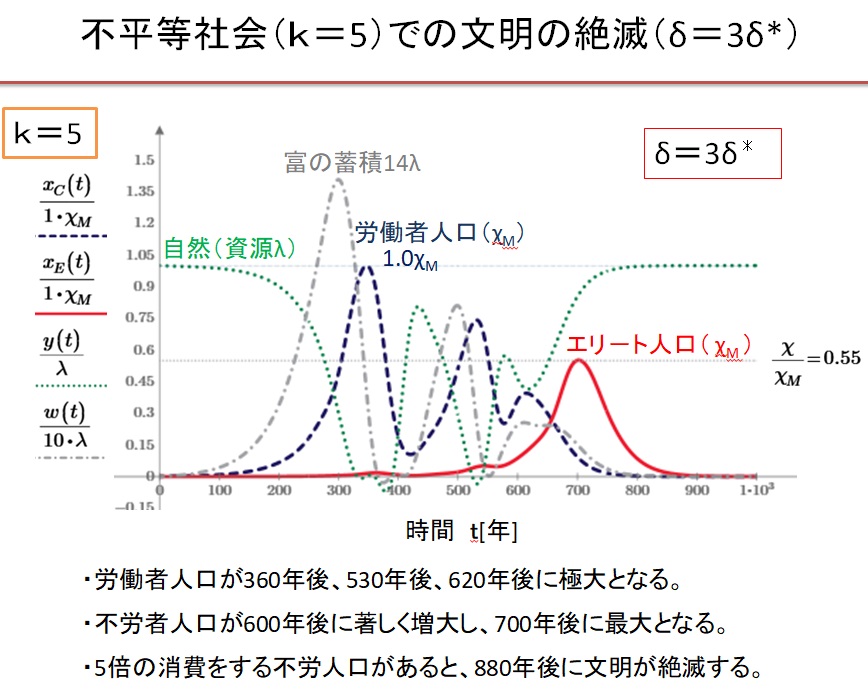
5.不平等社会での文明の絶滅
少数の不労者が一般人の5倍消費している不平等社会では、持続可能な社会が形成されず、文明は崩壊します。
(1)k=5の不平等社会でδ=1.0・δ*の場合
k=5の不平等社会において、Equitable社会で最大人口が達成できる収奪率δ*で収奪すると、文明は崩壊します。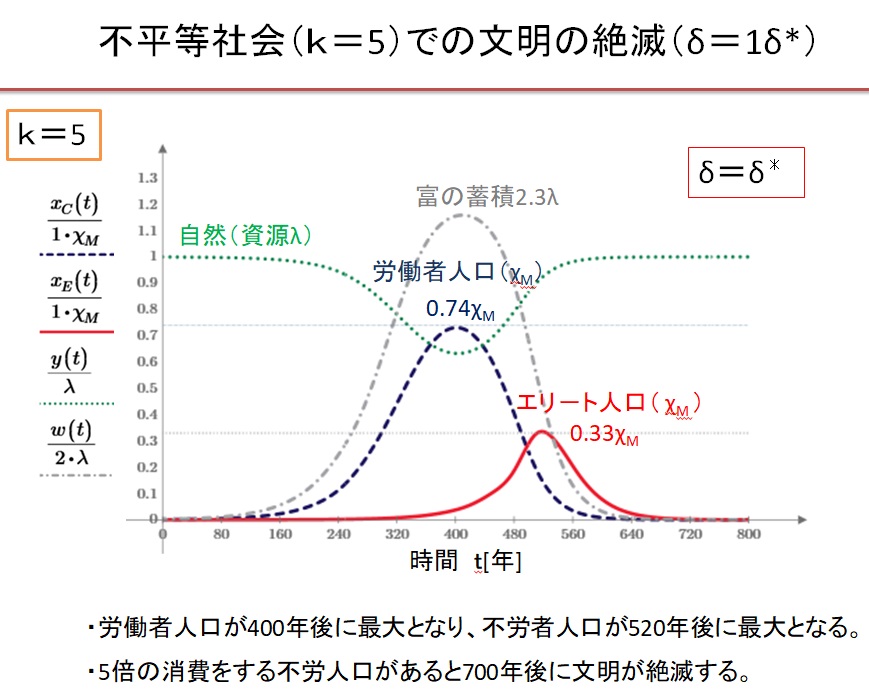
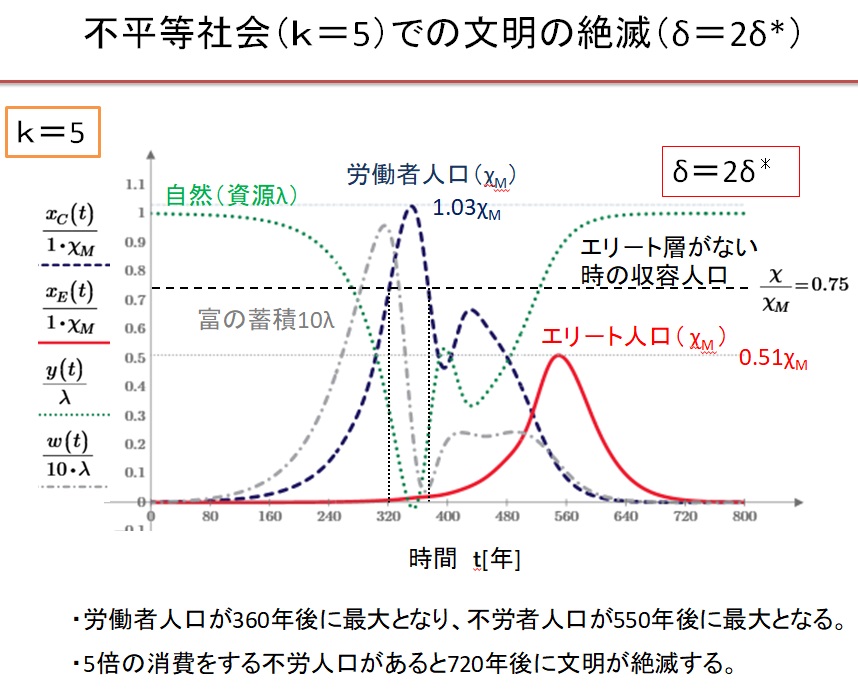
(2)k=5の不平等社会でδ=2.0・δ*の場合
(3)k=5の不平等社会でδ=3.0・δ*の場合
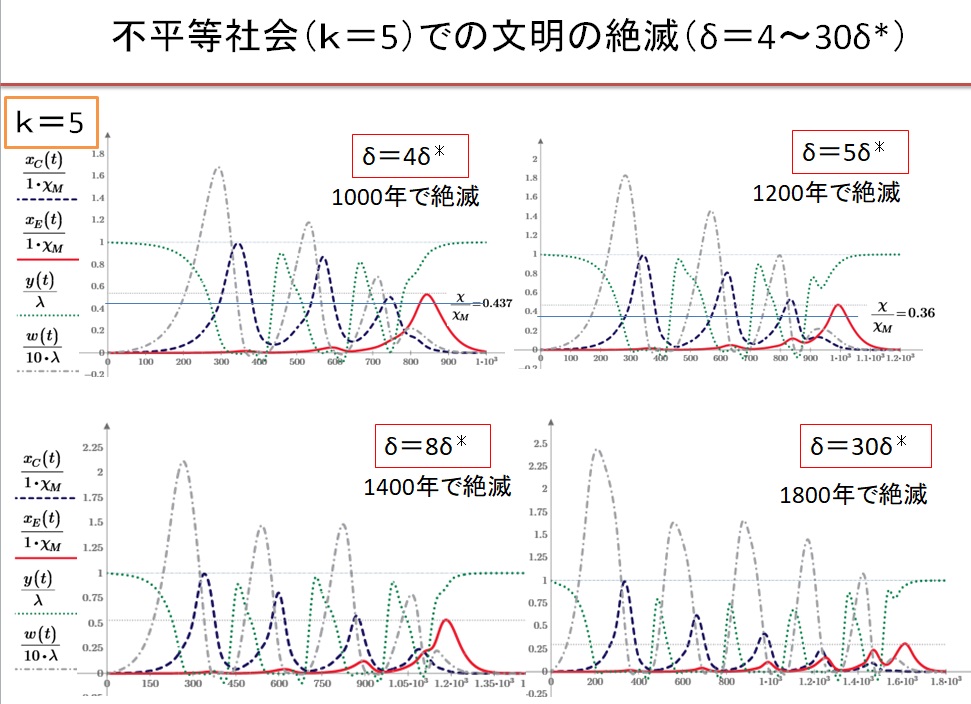
(4)k=5の不平等社会 δ=4~30・δ*の場合
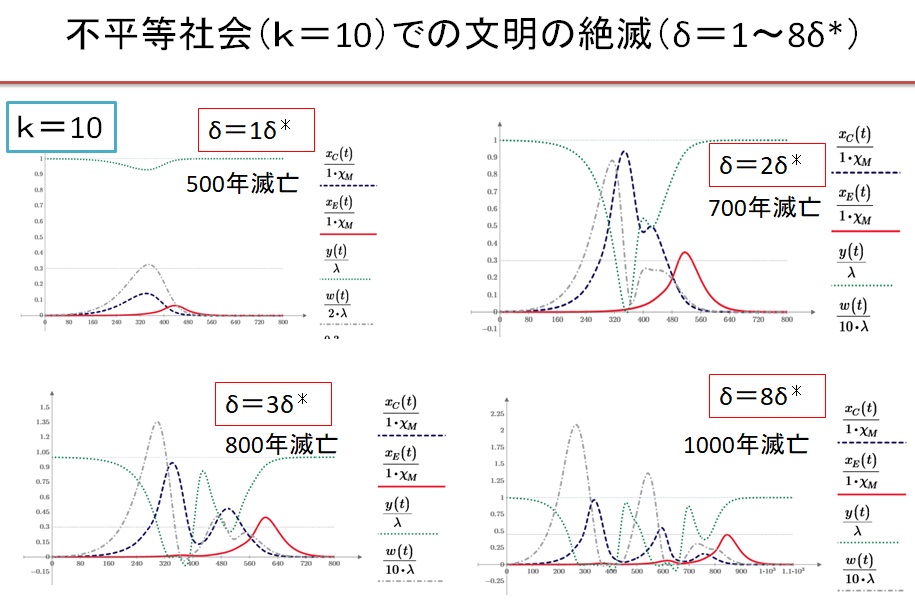
(5)k=10の不平等社会 δ=1~8・δ*の場合
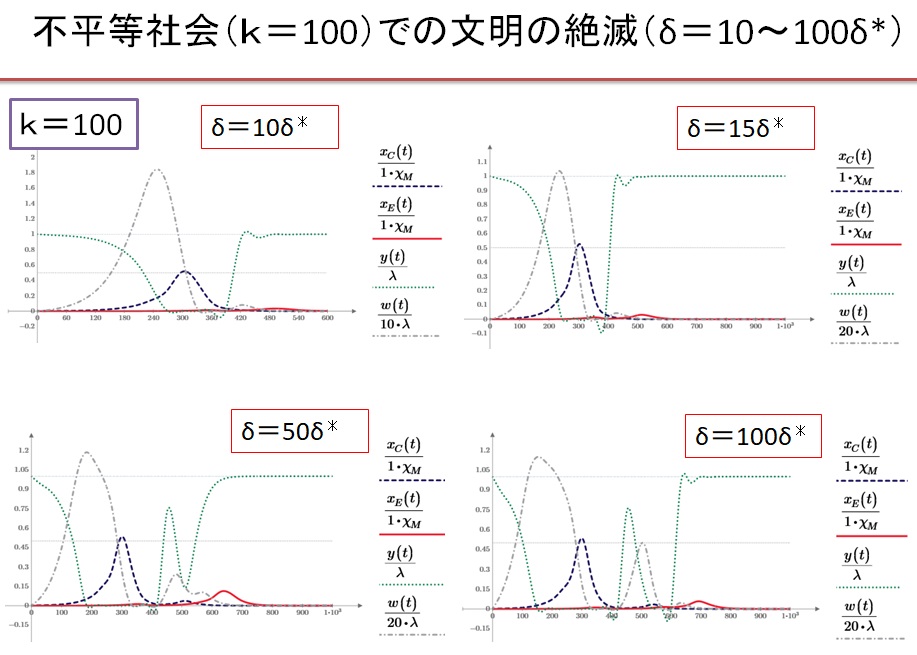
(6)k=100の超不平等社会 δ=10~100・δ*の場合
(7)k=100、δ=15・δ*の超不平等社会 初期人口依存性
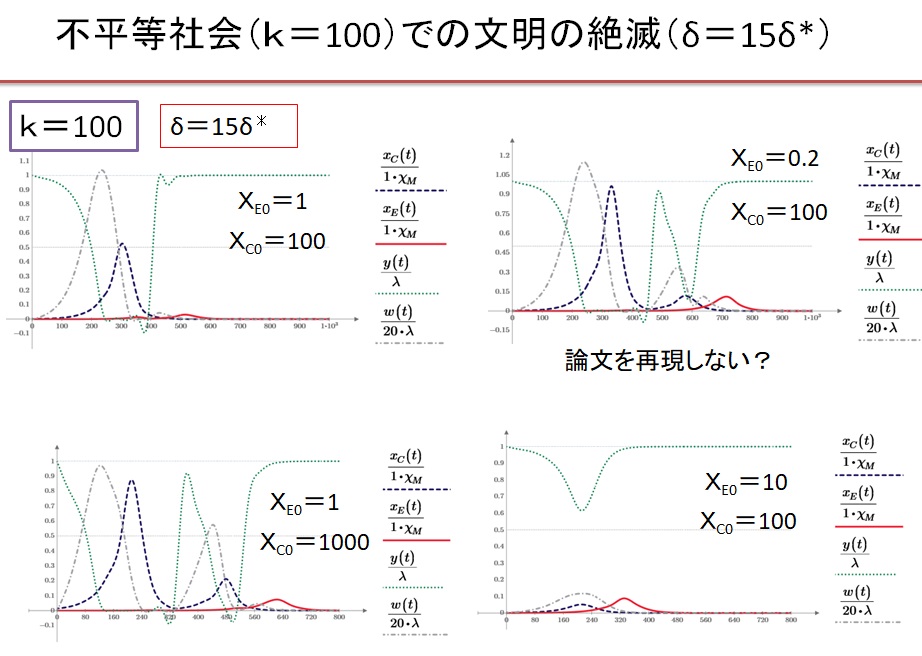
(8)k=10の超不平等社会 δ=1.2・δ*の場合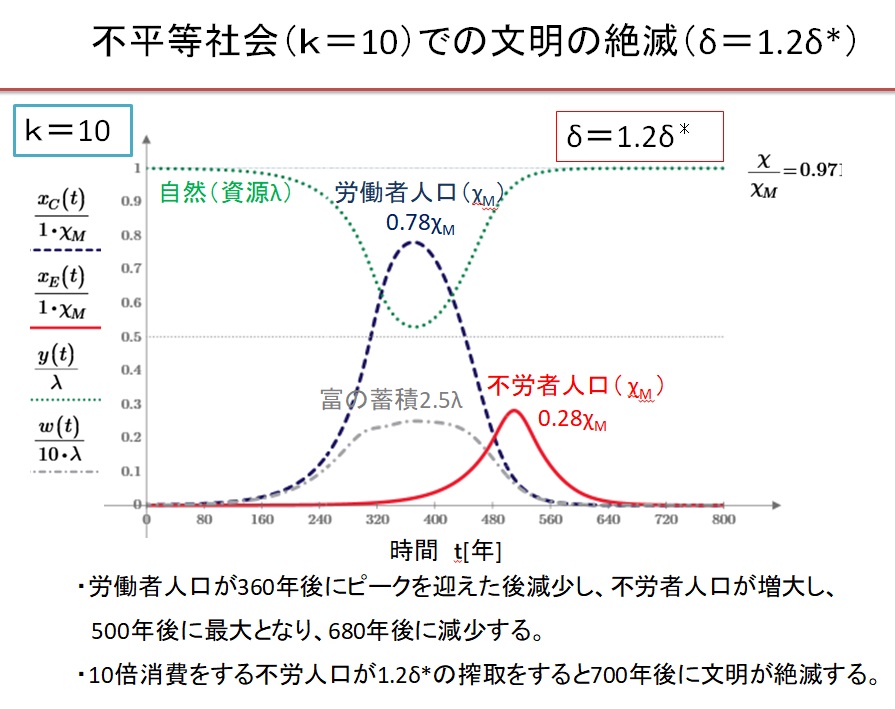
(9)k=100の超不平等社会 δ=15・δ*の場合
NHKスペシャルで紹介された上記条件の計算結果をほぼ再現した。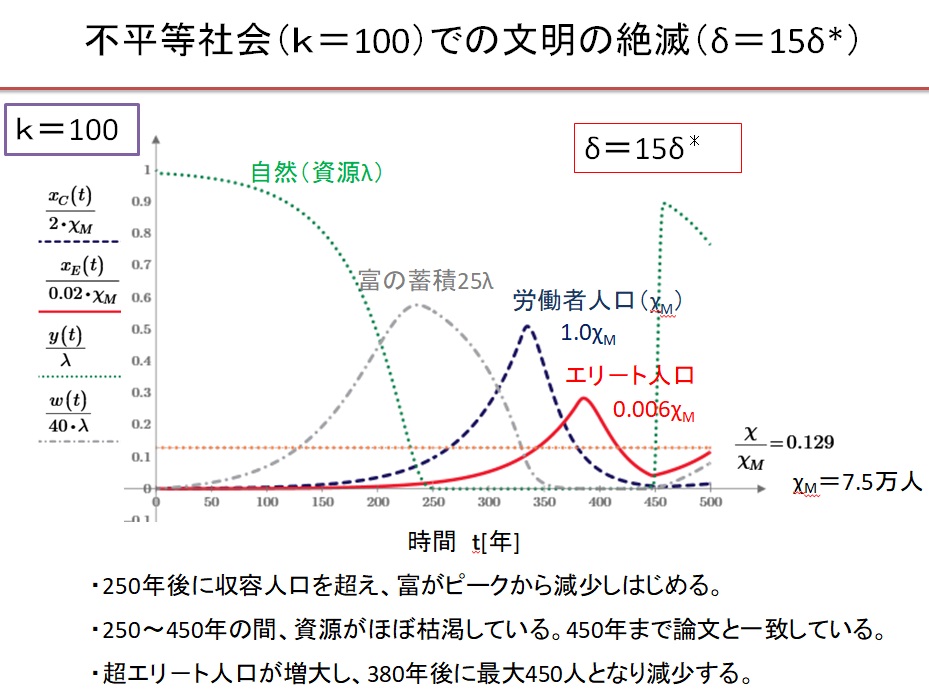
(10)NHKスペシャルで紹介された上記条件の計算結果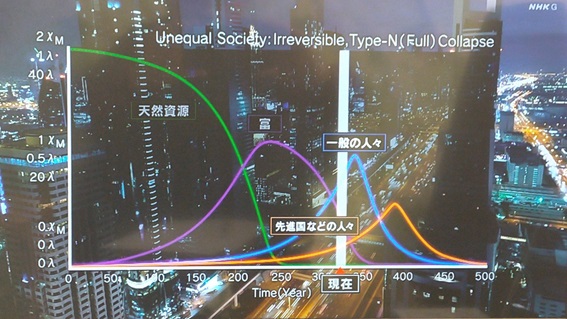
6.結果とまとめ

7.モデルの限界
このモデルでは、人口が減少し絶滅しそうになっても、エリ-トの消費量は一般人のk倍を維持していると仮定しています。現実には、人類が絶滅しそうになったら、エリ-トの消費量は一般人と同等になっていくのではないかと思われます。完全に消費量が同等になれば、不平等社会からEquitable社会へ移行し、持続可能な状態が実現します。しかしEquitable社会から不平等社会に逆戻りしたり、不平等が少しでも残れば、文明は絶滅する可能性が高いと思われます。
また文明が継続し技術革新によって死亡率が減少する可能性は考慮されていません。今後の課題としては、そうした修正をいれたモデルの検討が考えられます。
なおこのモデルでは、文明の絶滅原因を資源や富の減少に限定しています。実際は火山の噴火で生き埋めになったり、干ばつで水源が枯渇したり、大規模な洪水や地震などの災害や疫病の蔓延で文明が崩壊する可能性もあります。
新型コロナウイルス(SARS-CoV-2)の受容体はACE2である事は広く知られていました。最近の論文ではACE2の他にCD4、CD147、GRP78の受容体も報告されています。
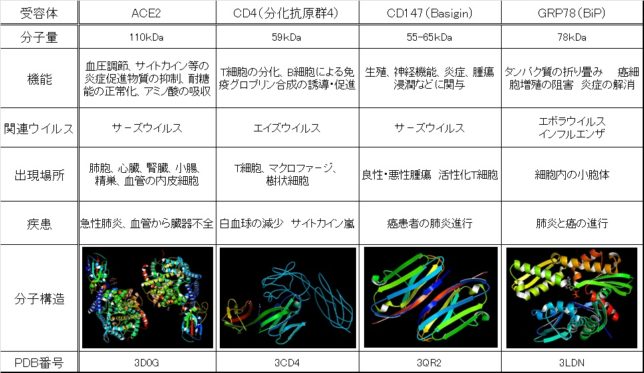
ACE2は十二指腸、小腸、胆嚢、腎臓、精巣に高く発現しており、副腎、結腸、直腸、精嚢に低く発現しています。ACE2は血圧調節以外に、サイトカイン等の炎症促進物質の抑制、耐糖能の正常化、腸でのアミノ酸の吸収などの働きをしています。高血圧、血管炎症、高血糖、胃腸障害などの基礎疾患がある人はACE2受容体の出現が増加するために、新型コロナウイルスに感染しやすくなると考えられます。新型コロナウイルスにより、ACE2が極度に減少すると、高血圧、急性肺不全、血管の炎症亢進、耐糖性能の悪化、腸障害などの症状がでて、重篤化すると考えられます。このACE2受容体は、炎症を起こし激しい咳き込みを繰り返した時などに多く肺胞細胞表面に発現すると言われています。そのためリコンビナント(組み換え)ACE2の投与実験がマウスにおいて研究されています。大量生産しやすいバクテリア由来ACE2様酵素も症状への効果が期待されています。
新型コロナウイルスが上気道(鼻腔、咽頭、喉頭)に留まっているうちは、ただの軽い風邪です。上気道のACE2密度は低いと言われています。この間は、自然免疫を担うNK細胞(ナチュラルキラー細胞)とウイルスの戦いにより、一週間程度の発熱が続くと考えられています。ウイルスに感染すると発熱するのは、体温を上げてウイルスの活性を低下させるために、血管を収縮させて放熱を下げるような対抗システムが備わっているためです。
肺胞の毛細血管の内皮細胞がACE2受容体を介してウイルスに感染すると、その情報が伝達されて毛細血管の平滑筋細胞が収縮し、体温が上がります。体温が上がると白血球が活性の低下したウイルスを活発に貪食します。ウイルスが消滅すると、体温が回復します。
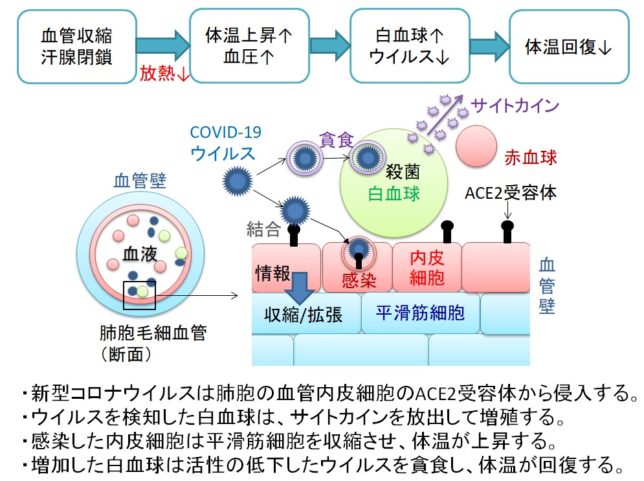
1918年に流行したインフルエンザでは、感染した米軍兵士がアスピリンを大量に服薬して、症状を重篤化させました。アスピリンは血管を拡張させて頭痛を除去する薬です。血管が拡張すると肺胞に水が溜まり、細菌感染を併発して、多くの人が亡くなったと言われています。流行の第二波ではインフルエンザの変性も生じて、感染率が高くなったと言われています。
新型コロナウイルスが下気道(気管支、肺胞)に拡散すると、自覚症状なく急性肺炎になるので、感染が疑われたら肺のCT画像を撮る必要があります。
CD4(Cluster of Differentiation 4、分化酵素群4)はT細胞、マクロファ-ジ、樹状細胞などの免疫細胞に発現するタンパク質受容体です。CD4はT細胞の分化、B細胞による免疫グロブリン合成の誘導・促進の作用があります。CCD4はインタ-ロイキン(IL)によってTh1、TH2、Th17のヘルパ-T細胞に分化し、それらは各種のインタ-ロイキンやイエンタ-フェロンなどのサイトカインを放出し、細胞内の細菌、寄生虫、細胞外の細菌を攻撃します。しかしそれらのサイトカインが過剰に分泌されると、炎症性疾患、アレルギ-、自己免疫性疾患を生じさせる問題があります。エイズウイルスはCD4受容体から感染するので、免疫系を破壊していまいます。血液1μLあたりのCD4数が200個以下になったHIV陽性患者はAIDSと診断されます。免疫不全になったエイズ患者は弱い細菌に感染して肺炎などで死んでしまいます。新型コロナウイルスはCD4受容体から感染し得るので、免疫系の破壊やサイトカインの過剰分泌を引き起こし、病状が重篤化します。
CD147は、Basiginとも呼ばれるタンパク質受容体で、上皮細胞、がん細胞、活性化されたT細胞に発現しています。免疫が活発になるほどリンパ球が感染して減少するので、がん患者の肺炎が急速に進行します。
GRP78 (Glucose-Regulated Protein)あるいはBiP(Binding immunoglobulin Protein)は、インフルエンザ、エボラウイルスの受容体でもあります。GRP78は細胞内の小胞体に分布しており、タンパク質の折り畳み、癌細胞増殖の阻害、炎症の解消の機能を担っています。新型コロナウイルスがGRP78を介して感染すると、GRP78の機能が障害を受け、肺炎、臓器不全、癌などが進行します。
新型コロナウイルスは細胞のACE2受容体から侵入します。
表に新型コロナ肺炎とインフルエンザの特徴を比較して示します。
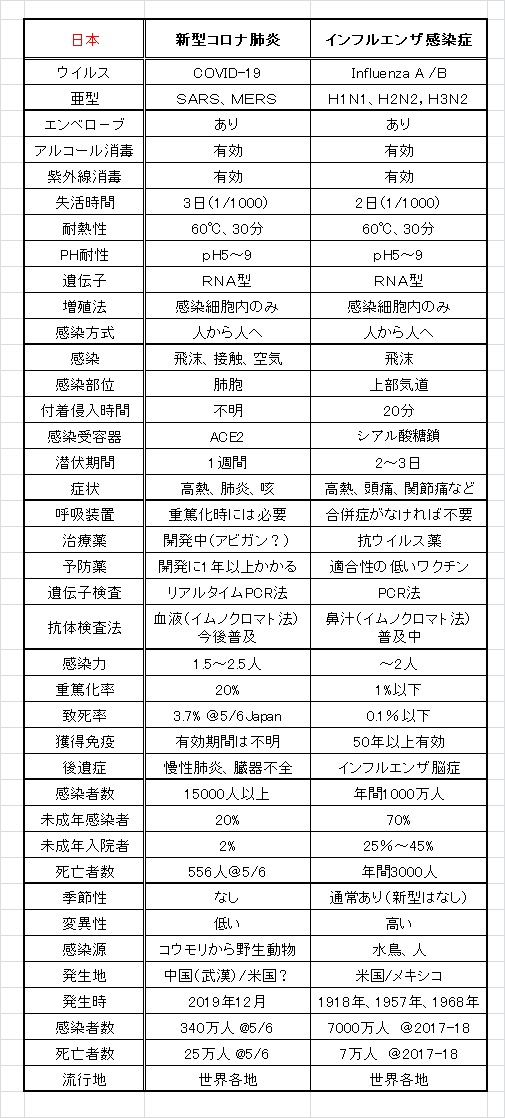
これら2つのウイルスはエンベロ-プをもつRNA型ウイルスです。エンベロープは、ウイルスのRNAを覆うカプシドを覆う脂質膜です。ウイルスが細胞外に出る際に細胞膜を被ったまま出芽することで獲得したものです。エンベロープには、ウイルス遺伝子がつくるエンベロープ・タンパク質が発現しています。エンベロープタンパク質は細胞側が持つレセプターに結合して、ウイルスが宿主細胞に吸着・侵入する役割を果たします。エンベロープは脂質から成るため、これらのウイルスはエタノールや石けんなどで処理すると容易に破壊できます。コロナ、インフルエンザの他に、風疹、日本脳炎、C型肝炎はエンベロ-プを持つRNA型ウイルスです。
天然痘や帯状疱疹やB型肝炎のウイルスはエンベロ-プを持つDNA型ウイルスです。帯状疱疹ウイルスは、水痘(Varicella)と帯状疱疹(Zoster)を引き起こすウイルスです。初感染時に水痘を引き起こし、治癒後に神経細胞周囲の外套細胞に潜伏しており、免疫力が低下するとウイルスが再び活性化して、皮膚に特徴のある帯状疱疹を引き起こします。ヘルペス用の抗ウイルス薬があります。
エンベロ-プを持たないDNAウイルスにアデノウイルスがあります。エンベロ-プを持たないRNAウイルスには、ノロウイルス、ロタウイルス、A型肝炎ウイルスがあります。エンベロ-プを持たないために、消毒が大変です。
アデノウイルスは直径80nmの正20面体のカプシドとそれに覆われた線状二本鎖DNAからなるウイルスです。アデノウイルスは感染性胃腸炎や風邪症候群を起こします。感染した場合、アデノウイルスは扁桃腺やリンパ節の中で増殖します。プール熱(咽頭結膜熱)とよばれ、高熱と微熱を繰り返す期間が4〜5日ほど続き、扁桃腺が腫れ、のどが痛みます。主要症状がなくなった後、2日間登校禁止となります。80%エタノールによる不活化時間は2分です。リネン類は、85℃10分間以上の洗濯、または水洗後に次亜塩素酸ナトリウム0.1%での消毒をします。
ノロウイルスによる集団感染は学校や養護施設などで散発的に発生しています。ノロウイルスは直径30-38nmの正二十面体のカプシドとそれに覆われたRNAからなるウイルスです。ノロウイルスは乾燥した状態でも、20℃で4週間以上感染力を保ちます。1968年、アメリカ合衆国オハイオ州ノーウォークの小学校において集団発生した急性胃腸炎患者の糞便から初めて検出されたので、ノーウォーク・ウイルスと命名され、ノロウイルスと呼ばれるようになりました。感染は経口感染で、腸から感染して増殖し、新しく複製されたウイルス粒子が腸管内に放出されます。10から100個程度の少数のウイルスが侵入しただけでも感染・発病が成立します。ノロウイルスはエタノールでは消毒できません。
インフルエンザは風邪より高熱が出て、関節痛などが生じる伝染病です。日本では毎年、新型インフルエンザが流行しており、ここ5年間は増加傾向にあります。日本のインフルエンザの感染者数は年間1000万人程度で、毎年3000人程度が死亡しています。つまり致死率は0.1%以下です。感染者の70%は未成年者であり、死亡者の殆どは免疫力の低い70歳以上の老人です。
日本の病院は1000万人のインフルエンザ患者に対応できるのだから、1.5万人の新型コロナ患者を受け入れるのはたやすいことだと考え、日本政府はコロナ感染拡大を誇張して報道させているのではないかと疑っている人がいます。しかし実際はそうではありません。
インフルエンザで入院する患者数は多い年で1万人近くになります。1週間経過後は徐々に回復して退院する人もでてくるので、ピーク時の在院者数は8000人(=9500人×0.85)程度だと推測できます。日本には感染者用の病床が12500床あるので、ピーク時の2月ごろは、毎年病床の40~65%がインフルエンザの患者さんで埋まることになります。これは決して余裕のある数字ではありません。実際は、病院数を削減しているので、高齢化社会になって増加するインフルエンザ患者を受け入れることは深刻な問題なのです。しかしインフルエンザの場合、学級閉鎖はあっても、サラリーマンに在宅勤務や主婦に外出制限をかけることはありませんでした。
社会がインフルエンザを許容しているのはいくつか理由があります。例えばインフルエンザの場合、
1) 重篤化率が1%、致死率が0.1%以下と低い。
2) 感染者の殆どが若者で、1週間程度で回復する。
3) 簡易検査薬や抗ウイルス薬があり、早期対応と症状軽減ができる。
4) 多くは季節性のもので、必ず収束する。
5) 感染後に獲得した免疫が持続するので、同じものには感染しなくなる。
6) 多くの人が免疫を獲得するので、感染が広がりにくくなる。
といった理由があるからです。インフルエンザは毎年流行するタイプが異なるので、完全に適合するワクチンが製造できません。完全に適合するワクチンを所有している人は、その新型インフルエンザウイルスを製造した人でしょう。そうなると流行は自然現象でなくなります。
それでは新型コロナ肺炎(COVID-19)はどうして社会的に許容されないのでしょうか? それは
1) 重篤化率が20%、致死率が4%以下と高い。
2)感染者の多くが高齢者や疾患保持者であり、重症化しやすい。
3)簡易検査薬や抗ウイルス薬が利用できず、早期対応と症状軽減ができない。
4)季節性がなく、潜伏期間が長く、感染が収束する保証がない。
5)感染後に獲得した免疫が持続するか不明である。
6)多くの人に免疫を獲得させられないので、感染が広がりやすい。
といった理由です。免疫の持続性が保証できれば、集団免疫獲得戦略を採用し、医者と病床数を確保できれば、社会的に許容する方向に進むかもしれません。しかし日本は医者の収入を守るために、医師数を厳しく制限しています。
現在の感染状況を見てみましょう。5月9日現在、東京都の新型コロナウイルス感染症対策サイト(https://stopcovid19.metro.tokyo.lg.jp/)によると、東京都の陽性者は4846人で、死亡者180人、退院者2152人、入院中2514人です。入院中の内訳は、軽症・中等症者2431人、重症者83人です。東京都の病床数は2000床なので、400人余りはホテルに隔離滞在していると思われます。陽性者の致死率は3.7%です。
ダイヤモンドプリンセス号の場合、3711人の乗客乗員のうち、712人が感染し、13人が死亡しました。感染率は19.2%、陽性者の致死率は1.8%でした。これは中国起因の感染で、現在東京都で感染しているウイルスは欧州起因のものだと考えられています。欧州起因のウイルスの致死率の方が2倍高いのかもしれません。
コロナ自粛の継続は、経済的な痛手ではありますが、日本企業にとって社員の健康管理や在宅勤務を取り入れた働き方を推進する良い機会になると思います。長距離通勤は、健康とエネルギと時間の浪費でした。行政を書面処理からオンライン処理に切り替えることで感染を避け、人員を削減できます。教育機関も遠隔教育を取り入れることで幅広い受講生を獲得できるでしょう。機械の身体を得て火星に移住する必要はありません。そんなことを学ぶために受験競争をするのはバカげています。勉強してマスクの一枚も満足に配れない総理大臣になっても仕方がないのです。教育内容は、専門バカを量産する教育から、豊かな生活と健康につながる教養教育に変化するでしょう。
豊かな生活は子どもに恵まれた生活です。産科医療を改革すれば、痛みなく障害のない健康な子どもを産んで育てられ、少子化問題が解決します。遠隔ワ-クにより自然環境のよい場所で子育てすることが可能になります。在宅ワ-クが中心になれば、地域社会に関わる機会が増え、高齢者になっても健康な生活が送れます。残業後にストレス解消にお酒を飲みすぎて体を悪くすることもなくなるでしょう。会社通勤や営業移動のストレスがなくなれば、健康に暮らせるのです。健康ならインフルエンザにかかっても、重篤化しません。早く治そうとして風邪薬を飲むこともなくなります。無駄な移動を無くせば、燃料の節約になり、交通渋滞や事故が減少し、大気汚染も防げるのです。コロナ自粛が日本の社会を改善するよい契機になることを願っています。
これから早急に答えを出していかなければならない難しい問題です。毎年日本では季節性のインフルエンザが流行し、多くの人が感染し、亡くなっています。しかし政府はインフルエンザの感染拡大防止のために外出自粛を要請することはありませんでした。その理由は、多くのインフルエンザの流行は自然現象であり、流行期間は集団免疫の獲得により3カ月程度で終了し、死亡者は老人が多く、社会に対する影響が限られているからです。
新型コロナ肺炎の場合、政府は7割~8割の接触削減を実現する外出自粛を要請しました。その理由は、国民の生命と生活を守るためです。具体的には
1) 新型コロナ肺炎の致死率が高く、放置すれば多くの死者が発生する。
2) 獲得免疫の持続期間が短ければ、流行が慢性化する。
3) 医療体制が崩壊すると、他疾患の患者が死亡する。
4) 長期の外出禁止政策により失業者が増大し、生活が困窮し治安が悪化する。
などの可能性があり、国民の生命と生活に与える影響が大きいからです。特に糖尿病や高血圧症や心疾患や腎疾患を有する人が新型ウイルスに感染すると重篤化します。
今後このような新型ウイルス感染症が発生するたびに国民の生命と生活が打撃を受けることになるでしょう。この問題が困難なのは、生命を守れば、生活が守れないというジレンマがあるからです。私たちはウイルス感染の拡大を早期に検出分析し、どの程度行動制限をかけるか、どの程度生活補償をするかを迅速に判断できる透明な体制を整えなければなりません。医療介護、小売り、清掃、配達員などの社会基盤維持に不可欠な労働者の生活健康保障も重要です。健康弱者の保護、健康格差の是正にも取り組むべきでしょう。
地震や津波などの自然災害とウイルス感染の流行が重なったら、避難や復興のために人が集まることで感染が拡大してしまします。こうした緊急事態に備え、対処していくことも課題になります。例えば隔離病床に使用できるホテル設計や遠隔医療は重要です。これからの日本の成功は、防災需要で経済を活性化させる仕組みづくりにかかっています。
日本政府は、4月7日に新型コロナウイルスの感染拡大を受け、改正新型インフルエンザ等対策特別措置法に基づく初の「緊急事態宣言」を発令しました。期間は1か月、対象地域は東京、埼玉、千葉、神奈川、大阪、兵庫、福岡の7都府県でした。4月17日に対象地区は7都府県から全都道府県に拡大されました。政府は感染拡大抑制のために、マスクの着用だけでなく、教育機関の授業中止、飲食店の営業自粛、観光客の観光自粛、公共交通の利用自粛、スポ-ツや文化活動の自粛など、全国規模の外出自粛を要請してきました。しかし医療機関は感染患者の受け入れのため病床数を拡大していますが、保健所による感染検査施設の設置は進まず、いくつかの病院では院内感染が発生しています。医療崩壊や介護崩壊を防止するために、日本国民はこれまで政府の自粛要請によく協力してきました。幸い5月6日の時点で感染者数は減少傾向を見せており、一部の地域の外出制限は解除されました。しかし東京都、北海道、石川県での非常事態は継続されており、国民の不安はまだ続いています。集団免疫が獲得できていないため、第二、第三の流行の可能性があるからです。
日中自宅で過ごしていると、新型コロナ関連のネット情報やテレビ番組が数多く報道されています。しかしウイルス感染症に対する科学的な理解を深める番組は殆どありません。マスコミは連日感染者数を報道していますが、具体的な治療内容は報道されていません。自粛要請で仕事を解雇されて、食費を減らさなければならない人も多く発生しています。多くの人は連休中に帰省することさえできなくなっています。地方自治体はすべての公園の駐車場を閉鎖したために、公園を自動車で利用する人は、健康のために公園で散歩することもできなくなりました。
このような社会的・経済的な停滞が長期化すると、国民の生活、健康、教育が疲弊していきます。政府は、この緊急事態にマスコミや支援給付金を利用して権力を強化し、国民を一方的に監視・管理する法律を成立させることもできます。そうなればたとえ感染者が減少しても一度成立した法律は元に戻らず、民主的社会が崩壊する恐れもあります。国家が新型ウイルスを合成し、ウイルス兵器と解毒剤を開発している可能性もあります。
私たちは、
1) 感染症を科学的に理解し、感染症を回避する。
2) 発熱した場合には、適切に行動する。
3)適切な食事や会話や運動をして、心身の健康維持に努める。
4)政府や自治体に合理的な政策を求め、私たちの自由と生活を守る。
5)ウイルスと共存してゆける社会を構築する。
といった新たな課題を実行していかなければなりません。この5つの課題について考えていきたいと思います。
コロナウイルスの感染が大きな社会問題になっています。感染抑制と経済損失の最小化を両立させるためには、感染率に応じた自宅待機率を実現しなければなりません。自宅待機率と感染率の間にはどのような関係があるのでしょうか?また回復にはどれくらいの時間がかかるのでしょうか?簡単な感染モデルで回復時間を見積もってみました。
感染者数をN[人] 、平均感染期間をT[時間]とすると、時間⊿tの間に増加する感染者数⊿Nは
・ ⊿N=-⊿t/T・N
となります。これを解くと
・ N=No・exp{-t/T}
となります。これは病院などでは感染者数が平均感染期間Tの間に治癒して減少することを意味しています。
次に感染者が自分は感染していると知らずに外出し、感染を拡大させる場合を考えます。自宅待機率をp、一人の感染者が単位時間に感染させる平均人数をK[人/人/時間]とすると、感染者数⊿Nは
・ ⊿N=K⊿t(1-p)N-⊿t/T・N
と表すことができます。(1-p)Nは外出している感染者の数で、第一項目は外出感染者が増加させる感染者数、第二項目は平均感染期間Tの間に治癒して減少する感染者数を表しています。これを整理すると
・ ⊿N/⊿t={K T(1-p)-1}・N/T
と書けます。これを解くと
・ N=No・exp[{K T(1-p)-1}t/T]
となります。つまり感染者数を抑制させる条件は、係数が負となる条件すなわち
・ K T(1-p)-1<0
となります。従って自宅待機率pが満たすべき条件は
・ p>1-1/KT
となります。KTは感染率すなわち一人の感染者が平均感染期間中に感染させる平均人数を表しています。例えば
・ KT=2.5 → p>60%
・ KT=5.0 → p>80%
となります。これまで政府は感染率2.5を想定し60%以上の自宅待機率を目指していましたが、感染率が2倍高ければ、感染抑制のためには80%以上の自宅待機率が必要になります。
ところで回復にはどれくらいの時間がかかるのでしょうか?感染者数が1/eになる回復時間(=37%に減る時間)は
・ Te=T/{1-K T(1-p)}
となります。平均感染時間Tは2週間程度だと仮定しましょう。下図に感染率KTが2.5(赤丸)、3.5(青ダイヤ)、5.0(緑四角)の場合の回復時間Teの自宅待機率p依存性のグラフを表示します。
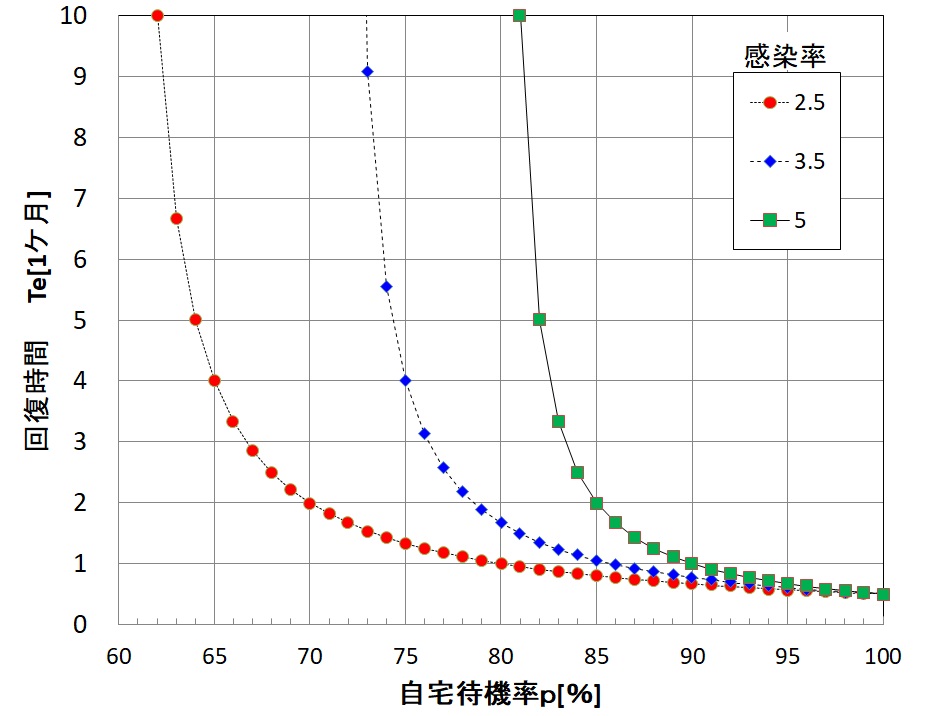
例えば感染率KT=2.5の場合(赤丸)はどうでしょうか?
自宅待機率p=61%であれば、
・ Te=T/{1-2.5・(1-0.61)}=T/0.025=40・T=20カ月
となります。自宅待機率が61%であれば、回復に1年8カ月かかることになり、来年の夏にオリンピックを開催することはできません。
自宅待機率p=65%であれば、平均感染時間Tは4カ月になります。つまり2月の時点で緊急事態宣言を発令していれば、6月には収束しているので、オリンピックは開催できたかもしれません。
自宅待機率p=80%であれば、
・ Te=T/{1-2.5・(1-0.80)}=2・T=1カ月
で回復します。政府が十分な休業補償をして、自宅待機率を80%に高めれば、回復時間が1カ月で済むということです。アメリカ政府はこのことをよく知っていたので、早期に莫大な休業補償に踏み切ったと考えられます。日本政府は十分な休業補償をしなかったので、回復時間が長引くことになります。全体の補償金額が増大し、このままでは国民は膨大な赤字国債を抱えることになるでしょう。
質問です。やる気のある人とない人が世の中にはいます。これが互いに迷惑を掛け合っている原因ですか?何故、同じ人間なのに分かれてるのでしょうか?(ayaさんより2020/4/5)
大変よい質問です。やる気のある人とない人がどういう割合で世の中にいるのか分かっています。パレ-トの法則をご存じでしょうか?パレートの法則とは、イタリアの経済学者ヴィルフレド・パレート氏が発見した冪乗則のことです。
例えば全世界の富の80%は20%の金持ちが所有している、会社の売上の8割は従業員の2割が生み出している、あるいは仕事の成果の8割は、費やした時間の2割の時間が生み出している、といった法則です。聞いたことはありませんか?
横軸に所有財産、縦軸に人数をとり、所有財産の分布図を書くことができます。金持ちになるほど人数は減っていきます。xを所有財産とすると、金持ちの人数は1/x^αに比例して減少します。べき指数αは問題によって変わりますが一定の値です。α=log45 ≈ 1.16とすると、80%-20%の法則が得られます。
つまり大部分の人は貧乏で、金持ちは一部の人なのです。金持ちだけ取り出しても、その中の20%の人はすごくお金持ちなのです。やる気は会社の売り上げだと考えればいいのです。横軸にやる気、縦軸に人数をとり、やる気の分布図を書くと同様のパレ-ト分布が得られます。やる気のある人は20%しかいないのです。
何故パレ-ト分布になるかは、今でも研究されています。一説には、全員がやる気をだすと、環境が激変したときに、社会が崩壊するかもしれないからです。例えばコロナウイルスが流行した環境下では、全員がやる気を出してお客さんに接触すると、全員が感染して、会社は崩壊します。実際は20%のやる気のある人が感染して退去しても、80%のやる気のない人が自宅で仕事を継続するので、会社は生き残れるのです。
一見やる気のない人がやる気のある人に迷惑をかけているように見えるのですが、緊急事態では、体力を温存していたやる気のない人たちが活躍するのです。といっても活躍するのはその中の20%の人だけです。迷惑をかけあっているのではなく、長期的に見て、助け合っているのです。やる気のない人が発生するのは、今の環境条件が決めているのです。人間に限らず、やる気のある働きアリも20%しかいません。つまり80%の人がやる気のない社会が、進化論的に生き残りやすいのです。だからやる気が出なくても自分や他人を無理に責める必要はないのです。
実は割れた窓ガラスの破片の大きさの分布や地震の頻度分布はきれいなパレ-ト分布になることが知られています。大きな地震は発生頻度が少ないのです。ガラスの破片を顕微鏡で拡大してみても、その分布は変わらないのです。そういう現象はスケ-ルフリ-現象と言われます。つまり現象の分布が拡大スケ-ルに依存しないということです。地震も岩盤という窓ガラスの崩壊現象だから、窓ガラスと同じ法則に従うのでしょう。所得や崩壊の分布がパレ-ト分布になるのは、両者が保存する富やエネルギの分配に関わるからなのかもしれません。
学校の入学試験の点数の分布は左右対称のガウス分布をしています。そうなるようにテストを作っているのです。しかし世の中の重要な分布は、ガウス分布ではなく、パレ-ト分布なのです。中央値は順位が中央の人の点数値です。ガウス分布では中央値は平均値と一致しますが、パレ-ト分布では中央値は平均値のずっと下です。順位が真ん中だからといって喜んではいられないということです。
将来の地震の規模や貧富の差は、ガウス分布で考えるより、ずっと大きいのです。貧乏人はそれに気づかないので、大災害に備えず、金持ちは妬まれずに安心しているのです。80%の人たちがどんな現象もガウス分布だと甘く見ているのは、パレ-トの法則なのかもしれません。
結局NADHを電子伝達系で用いる場合には、合計10H+がマトリクスから膜間腔へ輸送されます。FADH2の場合は合計6H+が膜間腔へ輸送されます。膜間腔のH+濃度と電位が高くなっているので、ミトコンドリア内膜を挟んだプロトン駆動力を利用してATP合成酵素がATPを合成します。
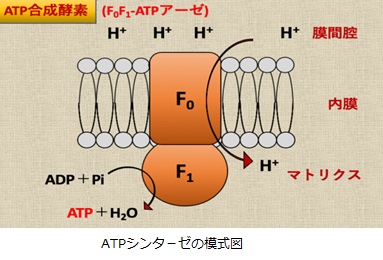
ATP合成酵素はF0サブユニットとF1サブユニットによる分子モ-タとして機能します。F0はミトコンドリア内膜に埋まっていて、F1はミトコンドリア・マトリクスに突き出た形で存在しています。F0モーターがH+の濃度勾配によるエネルギを使ってF1モーターを回すことによって、ATPを産生しています。3分子のH+がマトリクスへと輸送されるごとにATP1分子が合成されます。
しかし、実際にマトリクスにおいてATPを合成するためには、ATP合成の材料となるADPやリン酸Piをマトリクス内に取り込む必要があります。また、合成されたATPの大部分は細胞質で利用されるため、マトリクスから細胞質へと輸送される必要があります。
内膜を隔てたATP、ADP、Piの輸送
ミトコンドリア内膜を隔てたATP、ADP、 Piの輸送はアデニン・ヌクレオチド・トランスロカーゼとリン酸輸送体という2つの膜タンパク質によって行われています。アデニン・ヌクレオチド・トランスロカーゼは、ATP-ADP交換タンパク質のことで、細胞質のADP3-をミトコンドリア内へ、ミトコンドリア内のATP4-を細胞質へと対向輸送しています。この対向輸送では、プロトン勾配の電荷の差が用いられています。
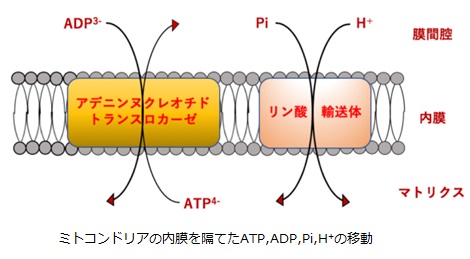 リン酸輸送体は、細胞質のPiをミトコンドリア内へと輸送するときにH+も同時にミトコンドリア内へと共輸送します。この共輸送ではH+の濃度差が用いられています。ちなみに、対向輸送とは、膜の内外で異なる物質を相互に逆方向に移動させる輸送のことで、共輸送とは、膜の片側から異なる物質を同方向に移動させる輸送のことをいいます。
リン酸輸送体は、細胞質のPiをミトコンドリア内へと輸送するときにH+も同時にミトコンドリア内へと共輸送します。この共輸送ではH+の濃度差が用いられています。ちなみに、対向輸送とは、膜の内外で異なる物質を相互に逆方向に移動させる輸送のことで、共輸送とは、膜の片側から異なる物質を同方向に移動させる輸送のことをいいます。
ミトコンドリアの外にATPを輸送し、マトリクスにADPを供給するためには、H+1個分のプロトン駆動力が用いられていました。ATP合成酵素は3H+で1個のATPを産出するので、細胞質内でATPを1分子増やすためには、4個分のH +が膜間腔からマトリクスに流入する必要があります。この数はNADHやFADH2が1分子あたりでどれくらいのATPを産生するかの指標となります。
細胞質内のATPを1分子増やすためには、4個分のH +が膜間腔からマトリクスに流入するということを踏まえると、NADH1分子あたり
・10[H+]/4[H+/ATP]=2.5ATP
FADH21分子あたり
・6[H+]/4[H+/ATP]=1.5ATP
が合成されることになります。
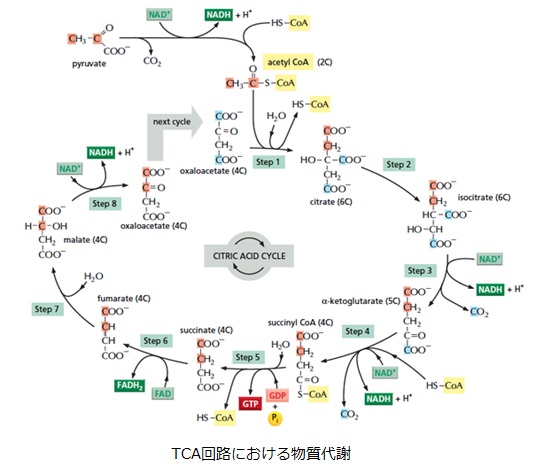
好気呼吸では、1分子のグルコースが「解糖系→ピルビン酸のアセチルCoAへの変換→クエン酸回路」という経路でATPやNADH、FADH2を生成していました。解糖の過程で2分子のATPと2分子のNADH、2ピルビン酸→2アセチルCoAの過程で2分子のNADH、クエン酸回路の過程で2分子のATP(GTP)と6分子のNADHと2分子のFADH2が生成されます。
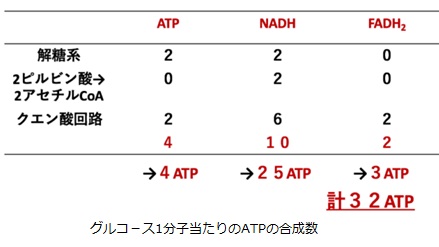
・4ATP+10NADH・2.5[ATP/NADH]+2FADH2・1.5[ATP/FADH2]=4+25+3=32ATP
結局1分子のグルコ-スは、嫌気的代謝では2分子のATPしか生成できませんが、好気的代謝では32分子ものATPを細胞外に生成できることが分かります。
細胞は糖を取り込み、ミトコンドリアで大量のATPを合成できることが分かりました。青森出身の安保先生によると、若い時は解糖系の瞬発力を主に使い、老年期になるとミトコンドリア系の持久力を主に使って活動するので、年齢ともに少食にしていった方が適正体重を保ちやすいそうです。
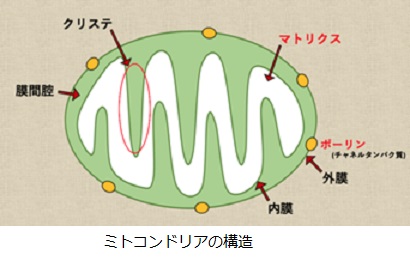
解糖系では1分子のグルコ-スは2分子のピルビン酸を生成するので2分子のNADHを生成します。さらに1分子のピルビン酸は、細胞内のミトコンドリアに送られ、ミトコンドリアのマトリックス内のTCA(tricarboxylic acid cycle)回路で3分子のNADHを発生させます。ミトコンドリアは外膜、膜間腔(まくかんこう)、内膜、マトリックスの2重膜構造を有しています。細胞によっては100~3000個ものミトコンドリアが含まれています。
運動してミトコンドリアが増えると同じ呼吸量でもATPの生産効率が高まるので、楽に走れるようになります。運動前は空腹にしておいて、最初に筋肉トレ-ニングをして汗をかいて有酸素運動状態に入ってから30分歩くだけでミトコンドリアは増加します。サウナの後に水風呂に入るとミトコンドリアは増加します。週末の2日間は摂取カロリを30%減らすのが有効です。日本医科大学の太田成男教授によると1日2時間の運動を1週間続けるだけでミトコンドリアは30%増加すると言われています。
TCA回路ではATP を直接作るのではなく、NADHやFADH2を作ります。さらにNADHやFADH2が呼吸鎖系でミトコンドリア内膜に水素イオンH+の濃度勾配を形成することにより、ATPを産生します。TCA回路は糖代謝だけでなく、アミノ酸代謝、尿素回路、糖新生など多くの代謝経路の仲立ちをしています。
TCA回路の全体反応は
・CH3-CO-S-CoA+3NAD ++FAD+2H2O+GDP+H3PO4
→ S-CoA+2CO2+3NADH+FADH2++2H++GTP
です。
NADHとFADH2はミトコンドリア内膜に埋め込まれた4つのたんぱく質複合体と反応してNAD +とFADに戻り、その際にミトコンドリアのマトリックスから膜間腔にH+を放出します。NADHは、解糖系で2分子、ピルビン酸脱水素酵素で2分子、TCA回路で6分子、合わせて10分子のATPを発生します。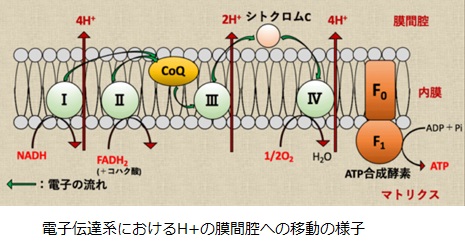 複合体ⅠでNADHはFMN(フラビン・モノヌクレオチド)と反応し、FMNに水素を渡します。FMNH2はFeSクラスタを介して、CoQ(ユビキノン)に水素を渡します。
複合体ⅠでNADHはFMN(フラビン・モノヌクレオチド)と反応し、FMNに水素を渡します。FMNH2はFeSクラスタを介して、CoQ(ユビキノン)に水素を渡します。
・NADH+H+ +FMN→ NAD++FMNH2
・CoQ+FMNH2→CoQH2+FMN
複合体Ⅱでは、コハク酸がフマル酸(2重結合あり)に変化するときには自由エネルギ変化が小さいのでFADが使われます。
・HOOC-CH2-CH2-COOH+FAD →HOOC-CH=CH-COOH+FADH2
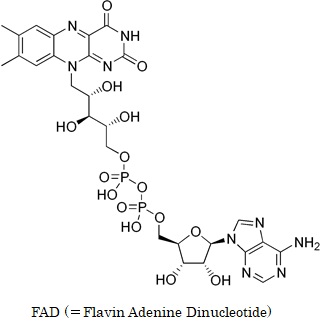
この反応で膜間腔に放出されるH+はありません。FAD (=Flavin Adenine Dinucleotide) はフラビン・アデニン・ジヌクレオチドの略語で、酸化還元反応における補酵素の一種です。FADの酸化還元電位は -219 mV で NAD 系より100mV程高く、開放エネルギが少なくNAD が使えないような反応で脱水素することができます。FADH2ではFADの左上の環が3つ並んだ部分の2つの酸素の二重結合がOH基になります。FADはADPにC5系炭素鎖と3環系のキノンが結合した構造をしています。
複合体Ⅲが行う電子伝達はQサイクルと呼ばれます。この反応では、まず、2分子のユビキノール(CoQH2)がユビキノン(CoQ)に変換される過程で4H+を膜間腔へと放出します。
・2CoQH2→ 2CoQ+4H++2e-+2e-
・CoQ+2H++2e-→CoQ H2
・Cyt(Fe3+)+2e-→Cyt(Fe2+)
なる反応が生じ、シトクロムcが還元されます。シトクロムcは膜間腔側にありヘム鉄(=鉄+ポルフィリン環)が含まれています。
複合体Ⅳは、シトクロムcオキシダーゼと呼ばれ、シトクロムc(Fe2+)を酸化して酸素を還元します。複合体 Ⅳが行う電子伝達の第一段階では、シトクロムc の電子がCuAに渡されます。その後、電子はヘムa→ヘムa3→CuBを経て、最終的に酸素(1/2O2)へと渡され、水(H2O)に変換されます。酸素分子の酸化還元電位は約 +810 mVであり、FAD よりはるかに電子を受け取りやすくなっています。
・Cyt(Fe2+)+2H++1/2O2 → Cyt(Fe3+)+H2O
このシトクロムcから酸素に電子が2個渡される過程で、2分子のH+がマトリクスから膜間腔へと輸送されます。複合体Ⅳにはヘム鉄(ヘムa)が多く含まれていますので、青酸カリがこのヘム鉄に配位すると、電子伝達系を阻害して、窒息してしまいます。
私たちは炭水化物を食べてエネルギ、すなわちATP(=Adenosine TriPhosphate)を生成して活動しています。1939年にEngelhardtらによって、筋収縮のタンパク質であるミオシンがATPを加水分解することが発見され、1942年にセント=ジェルジによってATPが筋収縮に関わるエネルギ源であることが解明されました。ATPはリボ-スの両側にアデニンと3リン酸が結合した構造をしています。
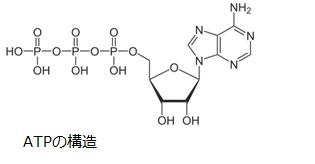 生体内では、ATPにリン酸1分子が離れたり結合したりすることで、エネルギの放出・貯蔵、あるいは物質の代謝・合成が行われています。ATPは加水分解によりエネルギを発生させます。酵素反応がATPの加水分解反応と共役することで、物質の代謝・合成が行われるのです。すべての真核生物がATPを直接利用しているため、ATPは生体のエネルギ通貨とも呼ばれています。
生体内では、ATPにリン酸1分子が離れたり結合したりすることで、エネルギの放出・貯蔵、あるいは物質の代謝・合成が行われています。ATPは加水分解によりエネルギを発生させます。酵素反応がATPの加水分解反応と共役することで、物質の代謝・合成が行われるのです。すべての真核生物がATPを直接利用しているため、ATPは生体のエネルギ通貨とも呼ばれています。
・ATP+H2O → ADP(アデノシン二リン酸)+ H3PO4(リン酸)
・ΔG°’ = −30.5 kJ/mol (=−7.3 kcal/mol) 標準自由エネルギ変化
細胞内では、ATP濃度はADPの10倍程高く、リン酸濃度も標準状態の1%以下であるため、細胞内の環境ではATPの加水分解に伴って放出される自由エネルギは−10〜−11 kcal/mol にもなります。
糖からATPはどのように産出されるのでしょうか?
炭水化物は胃腸で消化されて糖となります。糖は腸で吸収され血液と共に各細胞に送られ、細胞質内の解糖系で分解されてピルビン酸(CH3-CO-COOH)になります。嫌気的条件下ではピルビン酸は乳酸になります。好気的条件下ではピルビン酸は、CO2(=ピルビン酸のカルボキシル基に相当)を排出し、アセチル基(CH3CO-)になり、脱水素酵素においてNAD+を還元して、補酵素(HS-CoA)と不可逆的に反応し、
・CH3-CO-COOH+NAD+ → CH3-CO-S-CoA+CO2+NADH+H+
アセチルCoA(CH3-CO-S-CoA)を生成します。反応にはビタミンB1が必要です。これは不可逆反応なので、動物は脂肪酸から糖を合成できません。脊椎動物の細胞では糖から乳酸になるのは 4% 程度で、殆どは好気的にアセチルCoAを生成します。脂肪やたんぱく質も分解されてアセチル CoAとなってTCA 回路に入り、最終的に二酸化炭素 にまで酸化されます。過剰のアセチルCoAは中性脂肪を生成するため、アセチルCoAの代謝を抑制することで動脈硬化、高脂血症を防ぐことができます。アセチルCoAはADPに2つのペプチド結合を有する側鎖がついた構造をしています。
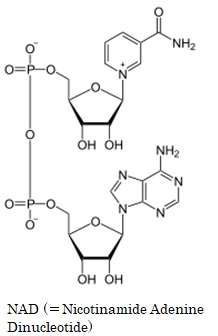
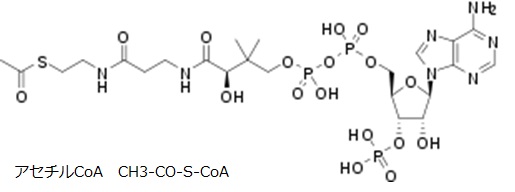 NAD (=Nicotinamide Adenine Dinucleotide) は ニコチンアミド・アデニン・ジヌクレオチド) と呼ばれる電子運搬体です。NADは2つのHを同時に引き抜き、自分がNADHになりつつ水素イオンH+を放出します。NADの酸化還元電位は‐320 mV と低く、異化代謝系で比較的大きなエネルギが解放される場合に、酵素反応に共役して脱水素反応を担います。NADはアデノシン・モノリン酸にニコチンアミド・リボ-ス・リン酸が酸素を介して結合した構造をしています。
NAD (=Nicotinamide Adenine Dinucleotide) は ニコチンアミド・アデニン・ジヌクレオチド) と呼ばれる電子運搬体です。NADは2つのHを同時に引き抜き、自分がNADHになりつつ水素イオンH+を放出します。NADの酸化還元電位は‐320 mV と低く、異化代謝系で比較的大きなエネルギが解放される場合に、酵素反応に共役して脱水素反応を担います。NADはアデノシン・モノリン酸にニコチンアミド・リボ-ス・リン酸が酸素を介して結合した構造をしています。
草食度物は球根(芋)や茎や葉を食べるので、グルコ-スが脳に供給されていると思うかもしれません。例えばウシなどの反芻動物は、細菌の力を利用して、前胃(ル-メン)の中で砕いた食物繊維やデンプンを酪酸やプロピオン酸に分解し、ルーメン壁から吸収することによって、エネルギの80%を得ています。酪酸はケトン体の前駆体であり、脳はケトン体をエネルギにすることができます。ケトン体とはヒドロキシ酪酸や、アセト酪酸などのことです。
ウシは、タンパク質を分解してできる尿素を唾液にまぜて、ル-メンに戻して細菌の窒素源にしています。増殖した細菌は4番目の胃で消化され、牛はエネルギとアミノ酸などの栄養を得ることができます。つまり草食動物は殆どデンプンをブドウ糖に分解していないので、草食動物の脳には危険なグルコ-スがあまり供給されていないのです。ウシも呼吸をするので、酸素を運搬する赤血球を養うために、ヒトの臍帯血の血糖値(35mg/100dL)程度のブドウ糖が必要です。しかしウシの血糖値は~50mg/100dL程度であり、ヒトの血糖値100 mg/100dLより低い値になっています。このように草食動物の脳には過剰なグルコ-スが来ないので、脳が大きくならなかったと考えられます。
私たちが持続的に生活していくには、様々な生物資源の管理と生態系の保護が必要です。生態系を豊かにしたのは、永い生物の進化の歴史です。生態系を理解するには進化の理解が欠かせません。ところで進化とは、どんなプロセスでしょうか?
進化は、変異、遺伝、選択の3つの過程によって成立しています。生物は、限られた食料と配偶者をめぐって生存競争をしています。遺伝子の突然変異によって、生存と繁殖に有利になり、適応力を高めた固体は、生存競争に勝って、多くの子孫を残します。その子孫は親の遺伝子を受け継ぎ、適応力の高い固体となります。
進化とは、適応力の高い固体に生じた遺伝子の変異が蓄積して、新たな種を生成することです。自然環境が適応力の高い生物の生存率や繁殖率を高めること自然選択といいます。生物が新しい形質を獲得していく機構を表したのが進化の方程式です。形質と言うのは、体長、眼の色、骨格、葉の形など、生物の色々な特徴のことです。

進化の方程式には色々ありますが、その中で有名なプライスの共分散方程式を紹介します。ジョ-ジ・プライス博士は生物群の世代交代による形質zの変化に関する共分散方程式
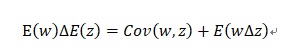
を発見しました。ここでE(w)は平均適応度、∆E(z)は形質の平均的な変化量、Cov(w,z)は適応度と形質の共分散、E(w∆z)は遺伝する形質変化を表しています。zは形質を何らかの形で数値化した変数です。この式は、形質の変化は形質に働く選択項と形質変化の遺伝項の和で書けることを示しています。変異は一定の割合で生じると仮定されています。以下に記号の定義を記します。
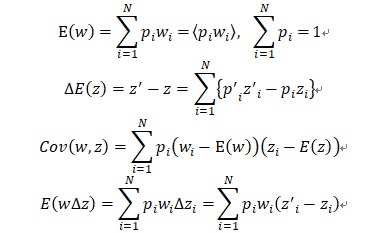 ここでwiは固体iの適応度、ziは固体iの形質zの値、piは固体iが形質ziを取る確率を示しています。さてプライスの共分散方程式を導出しましょう。形質の平均的な変化量∆E(z)は
ここでwiは固体iの適応度、ziは固体iの形質zの値、piは固体iが形質ziを取る確率を示しています。さてプライスの共分散方程式を導出しましょう。形質の平均的な変化量∆E(z)は
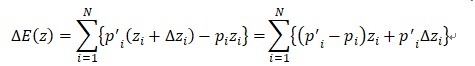
と書けます。適応度は子孫を残せる率に係る数値ですから、個体iの次世代の形質z’iを持つ確率p’iは
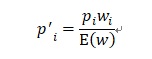 と考えられます。なぜならそれは前の世代の形質zを持つ確率piが適応度wiだけ増えるからです。E(w)で除することでp’iの確率保存
と考えられます。なぜならそれは前の世代の形質zを持つ確率piが適応度wiだけ増えるからです。E(w)で除することでp’iの確率保存
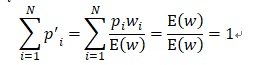
が成り立っています。p’iを上式に代入すると、 E(z)=0 として、
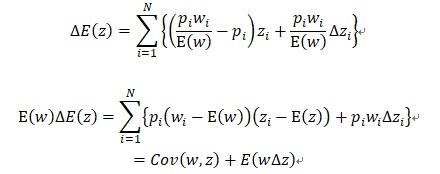
が得られます。Cov(w,z)は世代交代による適応度の変化による選択的な形質変化量、E(w∆z)は形質変化Δzが適応度wによって受け継がれる遺伝的な形質変化量を表しています。
上式の第二項を無視し、z=wとすると、
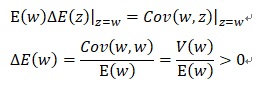
となり、平均適応度の変化∆E(w)は常に正となることが分かります。これは自然選択の第一法則と呼ばれています。そのためプライス方程式の第一項は自然選択項と呼ばれています。プライス方程式は、進化の基本方程式と呼ばれ、自然選択説に遺伝の効果を取り入れた特徴があります。この法則は進化学や生態学だけでなく生物資源管理にも適用できそうです。

ダーウィンは自然選択による進化論(1859年)の提唱者ですが、進化を論じる際に遺伝のことは全く考えていませんでした。遺伝学の祖メンデルは、30歳で物理化学を学ぶためにウィ-ンに留学し、気体反応の法則で有名なゲイリュサックや分子説で有名なアボガドロに会っています。そこで彼は、酸素ガスは2つの酸素原子が結合した分子であることなどを学びました。メンデルの法則(1865年)はAbといった2つの遺伝的要素で一つの形質を表現しています。このように遺伝学は分子説の影響を受けたと考えられています。
最近、テレビ番組では、地球温暖化の問題がよく取り上げられています。一般に地球温暖化などの環境問題や公共財の供給問題など、社会的ジレンマを含む問題が解決できないのは、全体にとって有益であっても、個人にとって不利益な行動は実現しにくいからです。しかし全体社会の中の小集団の支持者や利益を分析することで、政策を社会全体に浸透できる可能性があります。近年、社会的な政策は、個別地域に不利益があっても、全体的な政策支持の傾向があれば、実現可能かもしれないと考えられるようになりました。全体的な政策支持の傾向とは何でしょうか?具体的な数理モデルで考えてみましょう。


が得られます。ここでCovは小集団の人数niで重みづけた共分散です。〈xi 〉平均は小集団の人数niで重みづけた平均です。つまり共分散が正で、利得差を凌駕すれば、全体の政策Aの利得は政策Bより大きくなることが示されます。つまり法律や道徳的圧力がなくても、人々が徐々に政策Aを支持する可能性があります。但し小集団間でxiやuiのばらつきがある程度以上ないと、政策の誘導は難しくなる、というところが面白いですね。
プライス博士(George R. Price)は、無神論者で理論的に利他行動の可能性を追求してきたのですが、晩年はキリスト教に帰依して、ホ-ムレスに自分の財産を分け与えるなど利他行動を実践しました。1970年に進化生物学の基本方程式を発見し、1975年53歳のときにうつ病で自殺をしてしまいます。理由はお金がなくなってホームレスを助けることができなくなったから。追悼式に参列したのは、ビル・ハミルトン博士とジョン・メイナード=スミス博士と数人のホ-ムレスだけでした。オレン・ハ-マン教授が「THE PRICE OF ALTRUISM」(利他主義の対価)というプライス博士の伝記を書いています。最後に公式の証明を書いておきましょう。

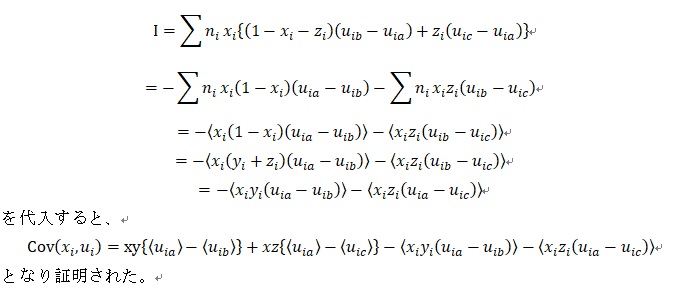
「森は海の恋人」という本があります。筆者は気仙沼湾の牡蠣漁師の畠山重篤さんです。畠山さんは、海の生物に栄養を供給している森の重要性に気づき、気仙沼に注ぎ込む大川流域の森林形成に尽力してしました。彼は講演会で、広葉樹の葉から生じたフルボ酸が岩から鉄を溶かし込むこと、フルボ酸鉄は川で運ばれ、海藻や貝を育むことを説明していました。
沈没船の周囲に海藻や魚が多いのは鉄が供給されるからだと言われています。しかし通常鉄は海水で酸化され沈殿してしまうので、海藻は鉄を利用できません。フルボ酸鉄は水溶性の2価の鉄Fe2+を供給するので、珪藻は鉄を吸収できます。鉄は珪藻細胞の呼吸系に必要なシトクロムの原料になります。海藻には川からくるフルボ酸鉄が必要であり、海藻を食する貝類や、海藻を住処とする魚を育てます。
ダムは、飲料水や産業水や電力確保のために建設されますが、森から供給される栄養を堰き止めてしまうため、下流の海は生き物の少ない海になってしまいます。不必要なダムは撤去しなければなりません。しかもダムの底には泥が溜まり、ダムの保水量は低下しています。日本の汚泥の蓄積量は15億m3、廃棄処理費用は14兆円以上と見積もられています。ダムの底泥を安易に山に引き上げると、大雨で流れて大規模な土砂崩れを引き起こしてしまいます。底泥の処理は頭の痛い問題です。
ダム底泥には有機栄養物とフルボ酸が豊富に含まれています。佐賀大学の兒玉宏樹教授らは、底泥からフルボ酸を取り出し、フルボ酸鉄を用いて海藻の養殖に取り組んでいます。福岡大学の渡辺亮一教授は、木屑と下水汚泥(または食品廃棄物)の発酵処理品と水処理剤として使用されているシリカ・鉄水溶液を混合し、2次発酵させ、安価なフルボ酸鉄シリカのペレット資材を開発しました(特開2018-143907 )。シリカは珪藻にケイ素を供給するために含まれています。そのペレット資材を袋に詰めて海のヘドロの干潟に設置したところ、珪藻が増え、それを食するアサリが顕著に増加し、ヘドロが分解することが確かめられました。

アサリは、濾過摂食者であり、成貝の濾水量は1個体で10L/日と多く、水質浄化と漁獲回復の双方を狙った干潟再生が期待されています。アサリの収量はかつての20万トンから1.4万トンに減少しています。
雄性不稔の遺伝子はミトコンドリアの環状DNAの中にあります。野生植物の場合、細胞の核内にある直鎖型DNAには花粉形成の回復遺伝子があり、ミトコンドリアの雄性不稔の遺伝子が働かないように抑制しています。
栽培植物では、突然変異によりミトコンドリアの雄性不稔遺伝子が無くなったために、核内の花粉形成回復遺伝子も不要になり喪失しました。そのため栽培植物は雄性可稔です。しかし野生植物と栽培植物の交配種の中には、野生植物に由来するミトコンドリアの雄性不稔遺伝子と、栽培植物に由来する花粉形成回復遺伝子のないDNAを有するものがあり、雄性不稔種が誕生しました。以下にこのことを詳しく説明しましょう。
Sを不稔(Sterility)遺伝子があるミトコンドリア遺伝子、F(Fertility)を不稔遺伝子がない可稔のミトコンドリアの遺伝子、RRを花粉形成回復遺伝子を有する核の優性遺伝子、rrを花粉形成回復遺伝子を有しない核の劣性遺伝子としましょう。そうすると野生種の遺伝型はS-RR、栽培種の遺伝型はF-rrと書けます。野生種と栽培種が偶然交配し、
・ S-RR(野生種)× F-rr(栽培種の花粉)→ S-Rr(可稔株)
S-Rrなる可稔株が誕生したと考えられます。さらにS-Rr同士の受精により
・ S-Rr(可稔株)×S-Rr(可稔株の花粉)→S-RR(野生)、 S-Rr(可稔)、S-rr(不稔)
の3種類の種が生まれました。このうち、S-rrは、核がミトコンドリアの不稔遺伝子の発現を抑制できないので、不稔になります。すなわち雄性不稔種の遺伝型はS-rrと書けます。
結局、歴史的に見ると、雄性不稔種にはミトコンドリアの異常があるという言い方は正しくありません。なぜなら元の野生種にはすでに雄性不稔遺伝子が含まれていたからです。むしろ野生種が突然変異して栽培種が出現したことが異常なことだったのです。栽培種と野生種から生まれた可稔株同士の交配によって、必然的に雄性不稔株が生じます。
雄性不稔種と同じ遺伝型をもつ栽培種の花粉を用いることで、
・ S-rr(雄性不稔種)× F-rr(栽培種の花粉)→ S-rr(雄性不稔種)
により雄性不稔種を増殖できます。種苗企業はこのようにしてF1種を生産していると思われます。稔性回復種F-RRを用いれば、
・ S-rr(雄性不稔種)× F-RR(稔性回復種の花粉)→ S-Rr(可稔種)
により、F1種子の可稔性を回復することもできます。つまり優性な花粉形成回復遺伝子Rが雄性不稔遺伝子Sを抑制するので、S-Rrは可稔種になります。
自分の花粉であろうと他の花粉であろうと、種さえできればミトコンドリアの遺伝子は確実に種に受け継がれていきます。雄蕊(おしべ)がなくても他の植物の花粉が雌蕊(めしべ)に付着すれば種はできます。ATPを生産するミトコンドリアには雄蕊をつくることは大きな負荷なのです。それより種子を多くつくる方がミトコンドリアの遺伝子を多く残せます。ミトコンドリアは元来別の生物であり、増殖する意思が強いため、ミトコンドリアは雄性不稔の遺伝子を作り上げたと考えられます。雄性不稔の遺伝子は、ATP合成の遺伝子に隣接しており、雄蕊をつくるときに、ATP合成を阻害するので、雄蕊が正常にできなくなります。
ところが他から花粉が得られない場合には、雄性不稔種は絶滅してしまいます。植物は核内の花粉形成の回復遺伝子によって我儘なミトコンドリアの雄性不稔の遺伝子が働かないように抑制しているのです。
実はコラ-ゲンの重合にはビタミンCが必要です。ビタミンCは、コラ-ゲンに含まれるプロリンとリシンの水酸化反応を触媒するFeを還元し、再利用する働きがあります。ビタミンCは還元できるHが2個もあり、強い抗酸化力を持ちますが、このような構造を持つ栄養素は珍しいのです。
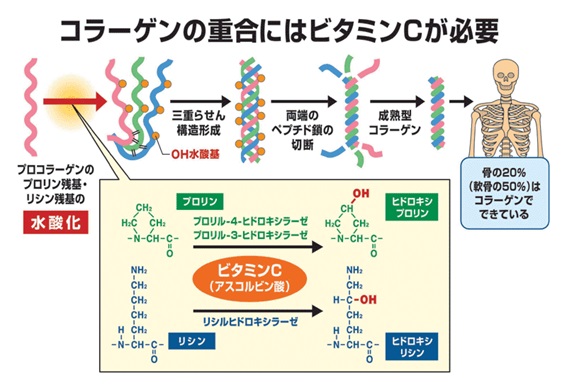
プロリンに水酸基が付加するとヒドロキシ・プロリン(Hyp)となります。α鎖のProとHyp間には水素結合があります。コラ-ゲン繊維のリシンと水酸化リシン間にはアルドール結合があります。ビタミンCがないと、Hypや水酸化リシンができないので、強固なコラ-ゲンが得られません(2013年、岸本)。ビタミンCが不足するとコラーゲンも不足し、血管や皮膚や骨が脆くなります。つまりビタミンCが不足すると、出血が止まらなくなる壊血病を引き起こすのは、コラーゲンが不足するからなのです。ビタミンCは、抗壊血病因子(anti-scorbutic factor)として発見されたことからアスコルビン酸(ascorbic acid)とも呼ばれます。酸化型はアセト・アスコルビン酸です。
ビタミンCは脂肪代謝にも関わっています。脂肪燃焼を促進するカルニチンはアミノ酸であるリシンとビタミンCから合成されます。ビタミンCはストレス抵抗ホルモンであるアドレナリンの分泌にも不可欠です。ヒトの場合、遺伝子欠損のため、ビタミンCは野菜や果物などの食物から摂取しなければなりません。
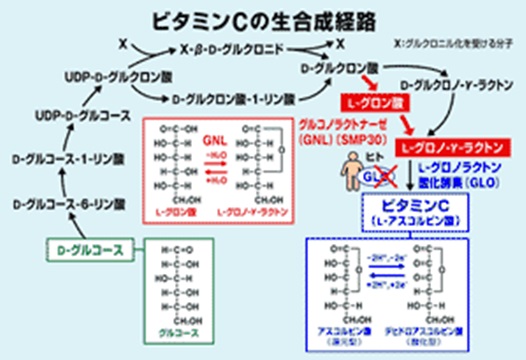
ちなみにヒトやモルモットは体内でビタミンCを合成できません。その理由は、ビタミンC生合成経路の最後に位置するGLO酵素(グロノ-γ-ラクトン酸化酵素)に遺伝子変異があるためです。GLO酵素を用いてグロノ-ラクトンからHを2個奪えばビタミンCが得られるのです。マウスはこのGLO酵素に遺伝子変異がないため、体内で充分量のビタミンCを合成できます。マウスとモルモットは違うのです。
コラ-ゲンは、基本的に人体の細胞を支える働きをしていますが、人体のあらゆる組織に存在し、多様な働きをしています。例えば、血糖調節、止血、骨代謝などもコラ-ゲンが関わっています。
止血のメカニズム(参考文献1)は複雑ですが、簡単に説明しましょう。止血を行う血小板の表面にはGP6という糖たんぱく質が突出しており、これがコラ-ゲンの受容体になっています。血管壁が損傷すると、I型やIII型コラ-ゲンのGly-Pro-Hypが、血管内に放出され、血小板のGP6受容体と結合します。血小板は刺激され、損傷部に凝縮し、血管を止血します。但しIV型コラーゲンは血管内皮基底膜の主要成分ですが、GP6を介した血小板凝集活性は認められません。
血管のコラ-ゲンが老化すると血管が固く脆くなります。血管全体の弾力性が失われるため、動脈硬化の原因となり、いわゆる高血圧・脳梗塞・心筋梗塞等のリスクを高めます。骨のコラ-ゲンが劣化すると骨折しやすくなります。プロリンは体内で合成されます。血管や骨をしなやかにするには、日頃からタンパク質を摂取して、運動をする必要があります。
参考文献1
コラーゲン結合タンパク質を介した生命プロセスの活性化機構 西田 紀貴
http://www.jbsoc.or.jp/seika/wp-content/uploads/2013/11/80-06-03.pdf
鮭のコラ-ゲンが変性しないようにするためです。コラ-ゲンに含まれるプロリンは環状の非必須アミノ酸です。プロリンは生体ではコラ-ゲン・タンパク質にのみ含まれています。ヒドロキシ・プロリンの含有量はヒトで9.5%、コイで7.5%、サケで5.4%です。ヒドロキシ・プロリンの含有量が小さいと、ラセンを保つ水素結合が減少するので、変性温度が下がります。コラ-ゲンの変性温度は、ヒトで42℃、コイで36℃、サケで21℃です。42℃はヒトの致死温度です。水温が25℃を超えると、サケは死にますが、コイは生きています。コラ-ゲンは生物の生存限界を決めています。ヒトが熱い風呂に長時間入るのは危険です。
それはイクチオステガ(Ichthyostega)だと言われています。イクチオステガは、約3.7億万年前(デボン紀最末期)に生息していた原始的四肢を持った魚です。Stegosは肋骨の覆いの意味です。脊椎動物が上陸するためには強い肋骨が必要だったのです。イクチオステガの化石はグリーンランドで発見されました。但し当時のグリーンランドは、赤道直下付近に位置していました。
体長は約1~1.5m。イクチオステガは四肢を使って移動し、尾でバランスを取っていました。少なくとも頭部を水の外に出すための強い前肢を持ち、頑丈な胸郭と背骨は日光浴の助けとなりました。幼魚時代には、優れた運動性により、水中の捕食者から陸上に逃れることができたと思われます。頑丈さゆえに体が重すぎること、尾に肉鰭類のような鰭を持っていること、後肢の先端には7本もの指があることなどから、殆どの時間を水中で過ごしていたと考えられています。3.5億年前の石炭紀前期に生息していたペデルペスが陸上を自由に移動できた最古の四肢動物だと言われています。

スクレロケファルスは、最初に海から陸に上がった開拓者「イクチオステガ」の直系の子孫にあたる両生類の仲間です。短足ですが、大きな体を十分支えることのできる足を持っていました。体長は1.5m程度あります。化石はドイツ(古生代 ペルム紀)で見つかったものです。

それはユ-ステノ・プテロン(Eusthenopteron:力強い鰭(ひれ)の意)だと言われています。ユーステノプテノンは3.7億年前(デボン紀)に現れた肺魚です。ユーステノプテノンはヒレの中に骨を持ち、肺で酸素呼吸をしていたので、湖沼のある湿地帯で生息することができました。食性は肉食性で、他の魚類を捕食していました。
1981年にアメリカのDonn Eric Rosen(1929―1986)らは、ユ-ステノ・プテロンは総鰭類よりも肺魚に近いと主張しました。ユ-ステノ・プテロンは植物の繁茂する河床に棲息していたため、密生した植物を対鰭でかき分けながら泳いでいたものと考えられています。体長は約30~120cm。体型はやや長い紡錘形で、上下に対称な幅広の尾ヒレがあります。
生息していた場所は海浜の潟湖などの気水域だったと推定されています。こうした場所は潮の満ち引きなどで環境の変化が著しく、水の流れが滞って酸欠状態に陥ることがあったと思われます。このことから、彼らは現在の肺魚と同じように肺呼吸をしていたと考えられます。さらには、鰭内部の骨や背骨、頭骨の構造が最古の両生類に近い特徴を示しています。

それはケイロレピス(Cheirolepis)だと言われています。ケイロレピスは3.9億年前(デボン紀初期)に現れた最初に背骨をもった魚です。ケイロレピスには2枚の胸ヒレと2枚の腹ヒレがあります。ケイロレピスの体長は約55cmです。ケイロレピスには顎(あご)と鋭い歯があり、自分の体長の2/3もの大きさの他の魚を食べられます。顎は、鰓(えら)を支える骨が稼働できる様に変化したものだと考えられています。ケイロレピスは背骨を持つことにより強い筋肉を発達させ、すばやく力強い泳ぎができたと考えられます。
淡水中のCa濃度は海水中の1~10%しかありません。ケイロレピスは体内に必要なCaなどのミネラルを骨に蓄積することで、川や湖などの淡水域で生息することができました。カルシウムは、神経の働きや心臓や筋肉が動くために必要です。骨にはMg、P、S、Znさらには鉄など、生命にとって必要なミネラルが海水と同程度の割合で含まれています。

軟骨は軟骨基質と軟骨細胞から成ります。軟骨基質の主成分は、コンドロイチン硫酸などのプロテオ・グリカンです。コンドロイチン硫酸は水和したNa+を引き付けるので、軟骨は豊富な水分を含んでいます。軟骨細胞は、軟骨基質の中の軟骨小腔と呼ばれる穴の中に入っています。血管は軟骨の中には侵入せず、軟骨細胞は、組織液を介した拡散によって酸素や養分を受け取り、不要物を排出しています。
軟骨は、軟骨基質の成分によって硝子軟骨、線維軟骨、弾性軟骨の3種類に分けられます。
硝子軟骨は、最も一般的に見られる軟骨で、関節面を覆う関節軟骨、気管を囲っている気管軟骨、胸郭の肋軟骨などがあります。均質、半透明で永久軟骨と呼ばれます。一方、哺乳動物の胎児は、全身の骨格が硝子軟骨として現れ、これが骨に置換されていきます。このような様式を軟骨性骨化といいます。
線維軟骨は、椎間板や関節半月などに見られます。軟骨基質にコラーゲンを多く含むのが特徴です。このため、軟骨としては固く、強い圧力に耐えることができます。靱帯や腱には軟骨成分は見られません。
弾性軟骨には、耳介軟骨や喉頭蓋の軟骨などに見られます。軟骨基質は、弾性線維を多く含むため、硝子軟骨や線維軟骨と比べ、柔軟でかつ弾力があります。
最古のサメは4.9億万年前のドリオドゥス(Doliodus problematicus)だと言われています。2017年にサメのような顎(あご)と胸鰭(むなびれ)の化石が見つかっています。サメの体は、殆ど軟骨でできているので、化石になり難いのです。
3.7億万年前(古生代デボン紀後期)の海にはクラドセラケ (Cladoselache) が生息していました。全長は約1.8メートルです。口は体の下側ではなく先端に位置していました。顎はあまり頑丈ではありません。体形が流線形であることから高速で遊泳していたと思われます。ちなみにサメの軟骨組織中には血管、神経、リンパ管はありません。
サメには腎臓がないと誤解している人が多いですが、サメやエイにも腎臓があります。サメの腎臓は、尿素を保持するために、尿素を再吸収します。そのためサメの腎臓は複雑な構造をしています。ヒトの腎臓は、尿を濃縮するために、水を再吸収します。水の再吸収時に尿素の浸透圧を利用しています。
初期の硬骨魚は弱者で、塩水域から淡水域に追いやられ、なんとか淡水に適応して生き延びました。現在の海の硬骨魚は、川から海に戻ってきたもので、海水を飲んで水を得ています。サメは海水を飲まずに海水から自然に水を得ています。サメは基本的には海水に適応していますが、ノコギリエイやオオメジロザメは、広塩性種と呼ばれ、海水と淡水の両環境に適応できます。
それはプテラスピス(Pteraspis)だと言われています。プテラスピス属は4.0億年前のシルル紀末に現れた腎臓をもった魚です。プテラスピスの体長は25cmほどです。プテラスピスには顎(あご)がありません。頭部は平たい三角形の外骨格で覆われ、三角形の左右の端と背中に棘が1本ずつ生えています。
海水に適用した魚が淡水に入ると体が膨れて破れてしまします。ステラスピスは体内に入り込んでくる水分を腎臓で体外に排出できたので、淡水域に生息することができました。恐らくこれによってプテラスピスは淡水域が苦手なオウムガイやサメから逃げることができたのでしょう。

尿素はサメの肝臓で合成されます。肝臓では尿素回路によりアンモニアから尿素を合成して補給します。尿素回路の律速酵素はカルバミルリン酸合成酵素です。尿素濃度が上昇すると、酵素活性が低下して、尿素の合成が止まります。サメの死後には尿素が分解されアンモニアになるので、臭います。しかし防腐効果があるので、内陸ではサメの肉が食されていました。サメの肝臓には肝油があり、肝油の浮力で、サメは遊泳しています。
鰓(えら)には、酸素を取り入れるために、大量の血液が流れています。海水と血液の間には僅かな細胞層で隔離されているため、サメは尿素を失いやすいのです。しかしサメの鰓の細胞膜はコレステロール含量が高いので、尿素の拡散透過性が小さくなっています。
尿素はタンパク質の変性剤として働くので、このままでは酵素活性が失われます。サメは、尿素とトリメチル・アミン・オキシド(TMAO)を2:1の割合に保つことで細胞内の酵素活性を正常に維持しています。サメの代謝系は尿素を前提として働いているので、淡水域のサメでもいくらか尿素を保持しています。
外界より体内塩分濃度が高いと、体内が脱水されてしまいます。魚の体内浸透圧の調整方法には3種類あります。クラゲや円口類のヌタウナギなどは、浸透圧順応型動物と呼ばれ、細胞内にグリシンやアラニンなどの中性アミノ酸を蓄積し、体内の浸透圧を外界と等しく保っています。
真骨魚は、浸透圧調節型動物と呼ばれ、過剰なNaCl はおもに鰓(えら)の塩類細胞から、Mg2+やCa2+、SO42-イオンは腎臓から少量の尿として排出されます。
サメやエイなどの軟骨魚やシーラカンスは、尿素浸透圧性動物と呼ばれ、体内に多量の尿素を蓄積することで、体内の浸透圧を外界と等しく保っています。サメの血漿(けっしょう)中のNaClは250~300mM(ミリモル/リットル)、尿素は400~450mMで、合わせて海水濃度700mMとなります。サメは鰓や直腸線からNaClを排出しています。Na+は、ATPを使ってK+と交換して、細胞外に排出されます。Cl-は濃度勾配や電荷反発を駆動力として排出されます。
最古の魚はアランダスピス(Arandaspis)だと言われています。アランダスピス属は4.6億年前、古生代オルドビス紀中期に出現しました。アランダスピスは体長15cm程度、頭は硬質で、鰭(ひれ)と顎(あご)はありませんでした。1959年にオーストラリアのアリススプリングスにて発見され、原住民のアボリジニのアランダ族から命名されました。
アランダスピスは海底付近をゆっくり泳いで藻類やプランクトンを泥ごと捕食していたようです。早く泳ぐ事が出来なかったため、巨大なオウムガイから逃げるように生活していたと考えられます。鰭がない魚がいたのですね。

同時期に生きていた似たような魚にアストラスピス(Astraspis)がいます。体長20cmで、名前は星の魚の意味です。体側にヤツメウナギのような8つのエラ穴が空いており、頭部の覆いが小さな五角形(星形)の骨片からできています。
自然の摂理と循環とは、「自然が、動物(捕食者)、植物(生産者)、菌類(分解者)が相互にバランスを保ち共存する摂理の元で、物質が循環再生産され、生物の持続的な生存と環境を実現している」ということです。菌類は種類が多く眼に見えないので理解が難しいものです。菌類を含めて、自然の循環を理解し、自然の循環に調和した暮らしを目指すことが、私たちの持続的生存を可能にします。
進化の過程で一年性の被子植物が多く出現しました。一年草は、個体の生存期間を短くすることで、遺伝子の多様性を増やしながら、速く増殖することに成功しました。これらの植物は、種を放出した後には、枯れて、土に還ります。大量の植物遺体が地表を覆えば、種には光が照射されないので発芽できなくなります。
しかし土壌菌は光を使わずに大量の植物遺体を分解します。そのおかげで植物は発芽できるのです。細胞質の糖や核酸は細菌によって比較的容易に行われますが、細胞壁は高分子の多糖類でできているので、分解は容易ではありません。前回は、細胞壁の分解は糸状菌(カビ)、放線菌(抗生物質を分泌する細菌)、担子菌(キノコ)などの多様な菌類の協力によって行なわれることを示しました。根の周囲のムシゲルなどの粘着性多糖類や分解されて残った有機物の一部はアルミニウムと結合し腐植になります。腐植は土壌の団粒化を促進します。腐植もまた徐々に分解されていきます。
増殖した菌類の遺体も菌類によって糖やアミノ酸に分解されます。有機物や菌遺体はさらに細菌によって、植物が吸収しやすい無機態の栄養素に変換されます。植物は、細胞の骨格となる炭素を光合成で得ています。しかしタンパク質や核酸や浸透圧調整に必要な窒素・リン酸・カリ(NPK)の殆どを主に無機物質の形で根から吸収しなければなりません。結局、植物や菌類の遺体の分解によって、遺体に含まれるNPKの栄養素やMg、Ca、Feなどのミネラルが土壌に供給され、それらは植物によって再び吸収され、再利用されます。
植物は、微生物の多様性を高めることで、病原菌から身を守っています。様々な菌がバランスよく生息している土壌は、活性の高い土と呼ばれ、病原菌のみが繁殖し難い状態になっています。植物は根から糖を分泌させることで、根の周りの菌叢のバランスを整えているのです。単一菌叢になると病気や連作障害が生じると考えられます。
植物は、根から糖やペプチドを出して、細菌を集め、グロマリンを放出し、土壌を団粒化し、菌類が棲息しやすい環境つくりをします。多くの植物は、菌根菌と共生し、根より細い菌糸を使って、細部の水分やリンやミネラルをより広範囲の土壌から得ることができます。また土壌の団粒化により、植物は自分自身に必要な水と空気が得られます。
団粒構造を著しく失った土は、降雨時には余剰水の涵水機能が働かなくなるので、畑の表面が川のようになってしまいます。そのような土地は雨があがったらすぐに乾いて、ひび割れを起こし、土埃を巻き上げます。適切な空気と水分がないので、微生物が住めなくなると、有機物は十分に分解できずに蓄積し、地下水を汚染するなどの問題を引き起こしてしまいます。
つまり土壌の団粒化は環境を保全します。団粒化により、降水が素早く深部に浸透するので、栄養素やミネラルが表面流出し難くなります。雑草や雑菌も栄養素やミネラルを土壌に保持する役割を果たしているのです。土壌の安易な耕起は、雑草や雑菌を殺し、土地の乾燥と荒廃をもたらします。団粒化は土壌生物によるものです。土壌の化学性と物理性を向上させには、まず土壌の生物性を向上させなければならないことに私たちは気づき始めたのです。
農に関する考え方
農に関する考え方を改めることは、環境を保全する唯一の道です。従来の農法のように土地を耕起して化学肥料を施肥し、優れた作物だけを収穫しようとすると、様々な問題を引き起こします。これからは自然の摂理と循環をよく理解し、菌相バランスの取れた健全な土壌を育成し、健全な土壌に作物を育成させなければなりません。山、海、川の自然環境と農とのつながりを見直さなくてはなりません。
持続的収穫
慣行農法では、畑の外部から種や肥料や農薬や農業機械など、大量の農業資材を投入しています。こうした農法は長期的には農地を疲弊させ、石油資源の枯渇後にも持続的に収穫していくことができません。
従来のように短期的な増収だけを目標にせず、これからは持続的な収穫を目標にします。そのためには、いきなり作物を栽培するのではなくて、食用にならないイネ科やマメ科の植物を栽培するなどして、時間をかけて健全な土壌を育成することも必要になります。健全な土壌や循環を取り戻した農地は、外から大量の肥料を与えなくても、持続的な収穫が期待できます。堆肥は増収のための手段ではなく、健全な土壌の育成のための過渡的な手段と考えます。堆肥の施肥の仕方も土壌環境を壊さない仕方に変えていきます。作物も単一ではなく多様な作物を栽培し、自然な環境を実現します。農業だけでなく林業や漁業も持続的収穫を目標にしなければなりません。
開かれた農
持続的収穫が実現できれば、農業はすべての人の生業になります。自分が食べる分を自分が作るのが基本になります。農作物を遠隔地に運ぶなどの様々な無駄がなくなります。家族や隣人との関わりも多くなるでしょう。自然から学び、自然と調和した生活は健康的です。土地に適した品種を選び、種採りして育てていくことで、災害に強く、安定した収穫が得られます。人々が広がって住めば、これまでの様々な問題が解決するでしょう。
堆肥の分解は、植物の細胞壁の成分を分解できる菌類が分解しやすい順番に分解します。堆肥は、落葉樹の枯葉、イネ科の枯草、米ぬかなどと水を混ぜて、保温して発酵させてつくります。米ぬかのリンPや窒素N分は菌体を作るのに使われます。N源として牛糞や鶏糞を使う人もいます。ときどき水をやり、切り返して均一に混ぜます。水をやらないと乾燥して、糸状菌が増殖せず、ヘミセルロ-スやペクチンの分解が進まないからです。ペクチンが分解すると、セルロ-スとリグニンが残ります。セルロ-スは放線菌が分解します。放線菌は60℃~70℃の高温に耐えられます。最後に残るリグニンは担子菌などが分解します。
分解の過程を調べるために、3種の菌類による植物の分解モデルを考えました。植物がペクチン、セルロ-ス、リグニンからなるとして、3つの分解菌(糸状菌、放線菌、担子菌)が分解する過程を表す数理モデルを立てました。
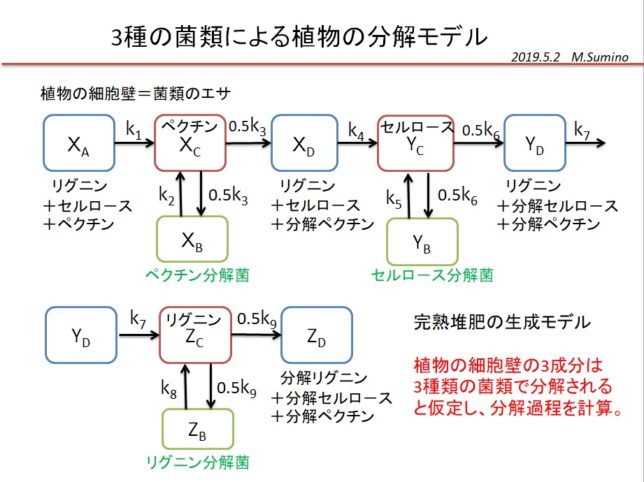
10元連立非線形方程式を市販のソルバで解いて、植物の分解過程を解釈しました。

植物を構成する成分は、ペクチン、セルロ-ス、リグニンの順番に分解され、十分時間が経つと菌類量は減少し、無害な堆肥が生成します。その様子を簡易モデルで実現することができました。
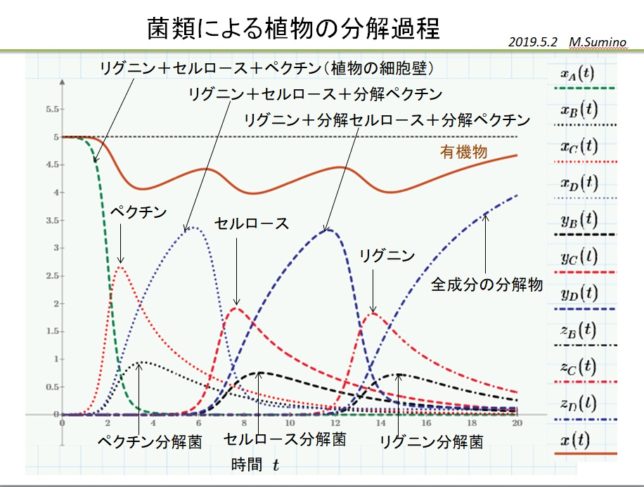
より複雑なモデルも考えられますが、結果は単純化したモデルと定性的に似ていました。
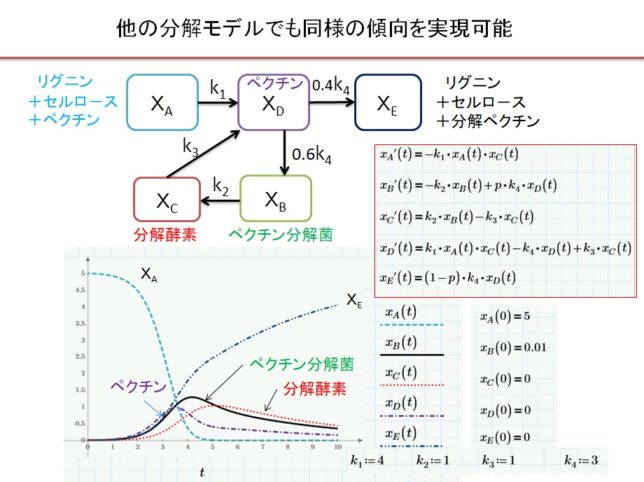
初期の菌数が少ないと増殖ピ-クを迎える時間が送れますが、定性的な形状は変わりません。分解が不十分な堆肥を施肥すると、残留糸状菌が作物に害を与えます。林の匂いがなくなるまで、堆肥を完熟させてから施肥します。
アルギン酸は、マンヌロン酸(M)とグルロン酸(G)という2種類のウロン酸が直鎖重合した構造を持つ多糖類です。アルギン酸の鎖状構造の中で、MとGはランダムに存在し、3種類のブロックを構成しながら共存しています。アルギン酸は、増粘剤、ゲル化剤、乳化剤、安定剤、麺質改良剤など食品の品質を向上する優れた機能を持っています。現在、アイスクリーム、ゼリー、パン、乳酸菌飲料、ドレッシング、即席麺、ビールなどさまざまな食品に利用されています。アルギン酸は、ペットフードや養殖魚の餌などに、結着剤や増粘剤として利用されています。
「落ちない口紅」にもアルギン酸が配合されており、唇の表面に被膜をつくって、口紅が移るのを防いでいます。アルギン酸カリウムは、アルギン酸のカルボキシル基にカリウムイオンが結合したかたちの塩です。その性質はアルギン酸ナトリウムと非常によく似ており、冷水や温水によく溶け、粘性のある水溶液をつくるとともに、Ca2+のような多価カチオンに接触すると瞬時にゲル化します。現在は歯科治療に用いる歯型取り剤(歯科印象剤)のゲル化剤として、国内外で広く利用されています。
ニトロゲナーゼは窒素に水素を付加する強力な還元剤ですから、酸素と容易に反応してしまいます。ニトロゲナーゼの金属クラスタは酸素に曝されると秒単位で速やかに分解されます。ニトロゲナーゼが失活しない酸素濃度は5~30 nM(モル濃度:M=mol/L)と非常に低いです。したがって、ニトロゲナーゼを駆動するにはO2を含まない嫌気環境が必要です。しかし生物が利用するATP生産の酸化的リン酸化のためには250 μMの酸素濃度が必要です。そのため窒素固定生物は様々な方法で嫌気環境を実現しています。嫌気性の窒素固定細菌は窒素固定に必要なATPを発酵など,酸素呼吸以外の系路によって生産しています。
根粒菌の場合
根粒菌はダイズの根など、レグヘモグロビンを含む根粒細胞に共生しています。レグヘモグロビンは酸素を強く捉え、酸素濃度に対する緩衝作用を有します。根粒の酸素拡散障壁を介した酸素濃度調節により酸素濃度は60nMとなり、レグヘモグロビンの酸素吸着作用、低酸素濃度での呼吸鎖の電子伝達を可能にする酸素高親和性のバクテロイド・ターミナルオキシダーゼによる酸素消費により、遊離酸素濃度は10nM程度になります。ちなみにヒトの血液の場合、遊離酸素の濃度は1μM程度です。
根粒菌の原形質膜の呼吸系はこの低濃度の遊離酸素を消費してATPを生産しています。そしてアゾトバクタと同様、このATPを利用して遊離酸素のない細胞内部に局在するニトロゲナーゼによって窒素固定を進行させています。この様に小さな細菌ではATP生産と窒素固定を、離れたところで進行させて、両立させています。
アゾトバクタの場合
アゾトバクタは、呼吸保護と呼ばれる細胞内酸素濃度を低く維持するための酸素消費速度の調節機構をもっています。呼吸保護には細胞表層に局在する5つのターミナルオキシダーゼによる酸素消費が大きく寄与します。それとともにアルギン酸が生合成され、細胞が覆われるアルギン酸の殻は細胞内の酸素を低くします。セルロース繊維のネットワ-クに水溶性のアルギン酸が裏打ちされると柔軟で酸素ガスを通さない膜が得られます。
細菌の表面膜の呼吸系で酸素を消費してATPを生産しています。アゾトバクタの細胞膜は酸素バリア膜であるアルギン酸膜で覆われています。外部から拡散してくる酸素は、細胞表面で全部消費されるため、細菌の内部には侵入しません。ニトロゲナーゼは酸素のない細胞の内部に局在し、ここで細胞の表面で生産されたATPを用いて窒素固定を行っています。
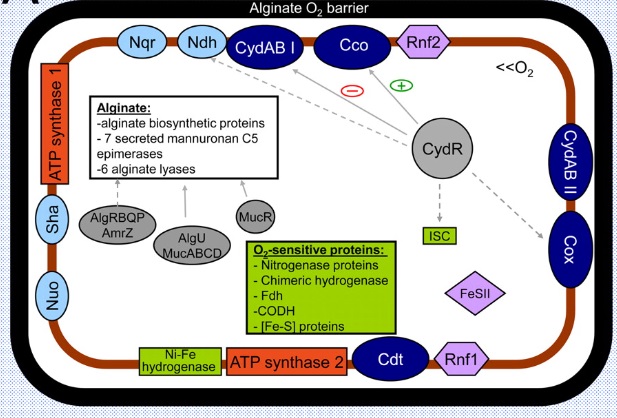
図1 アゾトバクタ属ビネランディの細胞の模式図
細胞はアルギン酸のバリア膜(黒)で覆われている。紺色のCydABⅠ、Cco、CydABⅡ、Cox、Cdtは5つのタ-ミナル酸化酵素、水色の4つの膜タンパク質Nuo、Sha、Nqr、NdhはNADHユビキノン酸素還元酵素、灰色のCydR、MucR、AlgUなどは制御タンパク質、赤色はATP合成酵素1とATP合成酵素2、紫色のFeSllとRnf1は呼吸保護に関わるタンパク質、黄緑の四角で囲われたものは、酸素暴露に敏感なタンパク質である。
参考文献:Joao C. Setubal, Virginia Bioinformatics Institute, JOURNAL OF BACTERIOLOGY, July 2009, p. 4534–4545,’Genome Sequence of Azotobacter vinelandii, an Obligate Aerobe Specialized To Support Diverse Anaerobic Metabolic Processes’
窒素固定シアノバクテリアの場合
光合成をするシアノバクテリアにも窒素固定をする種があります。光合成によりO2を発生しながら、酸素に弱いニトロゲナーゼを駆動するのは驚きです。ヘテロシストの膜成分は糖脂質です。細胞隔壁が外部からの気体拡散速度を調節することで細胞内酸素分圧を低減化しています。同時にヒドロゲナーゼ活性を高めることで環境中H2を酸化させて酸素を消費して細胞内酸素分圧を低下させています。糸状性シアノバクテリアは、環境中のC/Nが増加し窒素固定の必要性が高まった場合に、窒素固定専用の細胞(ヘテロシスト)にニトロゲナーゼを局在させ、光合成を行っている細胞からATPと 還元力をもらって、窒素固定を行っています。好気性の窒素固定菌は以上の様にして、酸素が窒素固定を阻害しないようにしています。窒素固定により生成したアンモニアは栄養細胞から供給されるグルタミン酸と反応しグルタミンへと変換され窒素源として栄養細胞に移送されます。
名古屋大学の藤田祐一教授は2018年6月にプレクトネマという窒素固定シアノバクテリアの20.8kbの窒素固定遺伝子のクラスタとCnfRという転写制御タンパク質を見つけ、その発現制御機構を解明しました。これらの遺伝子を、シネコシスティスという窒素固定の能力をもたないシアノバクテリアに導入して、窒素固定能を付与し、脱酸素試薬(ジチオナイト)の添加で、低いが有意なニトロゲナーゼ活性が検出しました。
フランキアの場合
フランキアは球状細胞ベシクルを形成することにより酸素の混入を防ぎ窒素固定を行います。ベシクルはホパノイド脂質の結晶からなる多重膜で覆われています。ベシクルの直径は4~5 μmで、ホパノイド脂質膜の厚さは50nm程度です。ベシクルは植物のミトコンドリアに取り囲まれており、これは酸素分圧を低下させる効果があると考えられています。
ニトロゲナ-ゼの反応中心は、[4Fe-4S]クラスタ、PクラスタとFeMo-coから成ります。Mo型ニトロゲナーゼは、容易に分離する2つのコンポーネントFe タンパク質(NifH二量体)とMoFe タンパク質(NifD-NifKヘテロ4量体)から構成されています。Nifは窒素固定Nitrogen fixationの省略記号です。NifHというのは窒素固定に関わるFeタンパク質をコ-ドしている遺伝子Hの名前です。Fe タンパク質は、ATPを加水分解してエネルギを得て、窒素の還元に必要とされる電子を送り出します。MoFe タンパク質は、Fe タンパク質から送られてきた電子を使って実際に窒素分子の還元を行います。
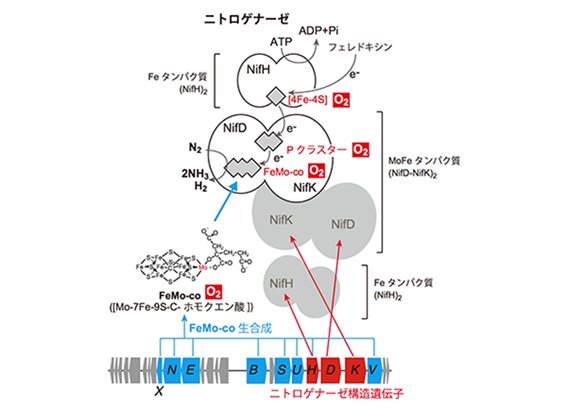
Fe タンパク質にある[4Fe-4S]クラスタは4つの鉄と4つの硫黄がキュバン(cubane)状(=立方体状)に集合したFe-Sクラスタです。MoFe タンパク質は、[8Fe-7S]構造のPクラスタとFeMo-coと呼ばれる[Mo-7Fe-9S-C-ホモクエン酸]の有機金属クラスタを含み、NifDとNifKの2つのサブユニットによるα2β2というヘテロ4量体構造をしています。[4Fe-4S]クラスタから送られてきた電子は、Pクラスタを経由してFeMo-co(フェモコ)に伝達され、FeMo-co上に結合した窒素分子を還元します。PクラスタからFeMo-coに電子を伝達すると、
となり、還元型のFeMo-coが得られます。さらに
となり、改めて生成されたFeMo-co(還元型)とN2H2が反応し
さらに改めて生成されたFeMo-co(還元型)とN2H4が反応し
によってアンモニアが生成されます。つまり還元型FeMo-coの力を3回使って、窒素からアンモニアが生成されます。
但しこれらの金属クラスタはすべて酸素によって速やかに破壊されてしまいます。また、ニトロゲナーゼの3つの構造遺伝子(NifH、NifD、NifK)に加え、FeMo-coの生合成には8つもの遺伝子が必要です。
窒素固定菌はニトロゲナーゼ(Nitrogenase)という酵素を使って、窒素分子をアンモニアへと変換します。炭化水素のC-H間の結合エネルギは約100 kcal/mol程度であるのに対して、N≡Nの三重結合のエネルギは225 kcal/molと高いので、これを還元的に開裂して2分子のアンモニアに変換することは大変なことです。Hoffman教授はニトロゲナーゼを“Everest of enzyme”と呼んだそうです(2009年)。ニトロゲナーゼは、モリブデンMoを含むMo型、バナジウムVを含むV型それに鉄Feのみを含むFe型の三種類あります。特に、土壌細菌のアゾトバクタ・ヴィネランディ(vinelandii)のMo型ニトロゲナーゼがモデル細菌となっています。ニトロゲナーゼは以下のような反応を触媒します。ここでPiはリン酸を表します。
電子はフェレドキシンなどの電子供与体から提供されます。ギブスエネルギの変化は
です。標準状態(PH7)にも関わらず大きな発熱を生じます。細胞での酵素反応中にATPは、Mg2+≡ATPの状態を取っています。つまりMg2+はATPの2つのリン酸の酸素イオンO-にキレ-ト結合しています。ATPの負電荷を覆うことで、Mg2+≡ATPが酵素反応の活性部位の疎水性の裂け目に結合できます。上記反応ではアンモニアに伴い水素が発生しています。ニトロゲナーゼは、ATPの加水分解と共役したプロトン還元の副反応
が含まれているからです。実際の生理状態においては16ATPではなく、20~30ATPが必要だとされています。ニトロゲナーゼは、反応特異性が低く、様々な窒素化合物や有機化合物を触媒できます。
生物による窒素固定量は年間5300万トンと言われています(植村誠次1977年)。その内、マメ科の根粒菌による窒素固定量は年間1400万トン、ハンノキ型根粒などの非マメ科によるものが500万トンで、全体の36%を占めています。それ以外の66%はアゾトバクタ-やシアノバクテリアなどの単独細菌によるものと考えられます。ちなみに人間が工業的に固定している窒素量は年間3000万トンです。
化学肥料は1ha当たり60kgの窒素分を投入します。マメ科の植物は68kg/haの窒素を固定し、アゾトバクタ-は50~280kg/haの窒素を固定するようです。アゾトバクタ-を有効利用できれば、窒素肥料は要らなくなりますね。
高橋英一(1982年)によると、窒素固定量は、ダイズ栽培土壌50~100kg/ha、クローバ栽培土壌100~200kg/ha、サトウキビ根圏土壌~60kg/ha、水田30kg/ha、アカウキクサ栽培池60~120kg/haです。
サトウキビが作物体に貯えた窒素の50%近く、サツマイモでは葉中窒素の40%近くが植物細胞に内生するエンドファイト細菌から供給されていると考えられています(涌井2003年)。すなわち、作物は肥料だけで成長するのではなく、窒素固定菌の働きが想像以上に大きいことが分かります。
窒素固定菌というとマメ科の根粒菌(Rhizobium)が有名ですが、他にも単体で窒素を固定するアゾトバクタ(Azotobacter)、クロストリジウム(Clostridium)、藍藻(diazotrophic cyanobacteria)がいます。またフランキア(Frankia)などの放線菌(Actinomycetales)は多くの樹木と共生し、空気中の窒素をアンモニアに変えて摂取しています。土壌の中のアゾトバクタが十分いれば、作物の幼苗は、窒素肥料を殆ど与えなくても、成長します。窒素肥料を与えてしまうと、アゾトバクタと作物との緩い共生関係は無くなってしまいます。牛糞堆肥などを使う有機農法から無肥料栽培に転換するには、土壌中の窒素成分を減らして、微生物の転換を図らなければなりません。
マメ科の根粒菌
マメ科植物の多くは根に粒状の根粒を形成し、そのなかに根粒菌が共生しています。根粒菌は植物から糖をもらい、植物に空気中の窒素を分解して得た窒素化合物やホルモンを供給しています。マメ科植物は世界各地に分布しており、現在450属、13,000種ほどが知られています。調査された2,000種のうち、根粒を形成しない種類が約10%ありました。
アルファルファ、クロ-バ、エンドウ豆、インゲン豆、ル-ビン、大豆、カウピ-(落花生)、ミヤコグサ、ダレヤ、イガマメ、ニセアカシア、イタチハギ、タチレンゲソウ、ムレスズメなど約20種類のマメ科の植物は、交互に根粒菌を交換しても根粒が形成されます。根粒菌は、土壌中では鞭毛のある小型の球菌ですが、共生する時は桿状大型化し、不規則な形態のバクテロイドになります。根粒組織中にはレグヘモグロビンという赤色の色素がみられます。根粒の寿命は多くは1年以内であって、結実するころから根粒の内容物は寄主植物に吸収され、根粒内の根粒菌は土中に放出されます。
根粒の形成には、好気的条件が必要です。また窒素肥料が多いと根粒が形成されません。リン酸は根粒形成に不可欠で、根粒形成を促進させます。微量元素の硼素(B)は根粒とバクテロイドの形成に、モリブデン(Mo)は窒素固定に必須の元素です。マメ科作物の種子に根粒菌を接種して根粒を形成させる人工接種の手法が開発されています。人工接種すると、無効菌が先に根に侵入する前に根粒が形成されるので、作物の収穫量と品質が向上するようです。
アゾトバクタ
アゾトとはイタリア語で窒素の意味です。クロオコッカムやビネランジは、シュ-ドモナス科アゾトバクタ属のグラム陰性の好気性細菌です。アゾトバクタ属の細菌は単体で自分のために窒素を空気中から取り入れ固定します。アゾトバクタは1gの炭水化物を消費して5~20mgの窒素を生産します。これは根粒菌の10%程度です。
アゾトバクタは、土壌中に広く分布し、中性付近で窒素固定を行うため、酸性土壌にはあまりいません。植物は根から糖を出し、アゾトバクタはアンモニウムを出して、お互いに緩い共生関係を保っています。特にアゾトバクタは光合成細菌と共生します。アゾトバクタは脂肪酸を提供し、光合成細菌は糖を提供します。そのため光合成細菌を施肥すると窒素固定量が増えます。アゾトバクタは大量の酸素を消費するので、環境が嫌気的になりがちですが、光合成細菌は酸素がなくてもATPを生産できるので、アゾトバクタと共生できるのです。
窒素固定細菌に関しては、アゾトバクター属以外に4属が知られています。
クロストリジウム
クロストリジウムは3~4μmサイズのグラム陽性の嫌気性細菌です。繊毛があり運動します。耐酸性があり、あらゆる土壌に分布しています。但し窒素固定力はアゾトバクタより弱いです。クロストリジウム属細菌は、SODやカタラーゼなどの活性酸素を無毒化する酵素を持たないため、酸素がある通常の環境下では不活化します。酸素存在下では、耐久性の高い芽胞を作って休眠することで、死滅を免れます。ボツリヌス菌や破傷風菌やウエルシュ菌はクロストリジウム属の細菌です。サ-モセラム菌は好熱性で酸素なしにセルロ-スを分解できるため、エタノール生産に利用されています。クロストリジウム属菌はガン細胞を選択的に攻撃することが知られており、その応用が研究されています。
窒素固定シアノバクテリア
単細胞・糸状体種のシアノバクテリアの半数は窒素固定の能力を持ちます。大部分は単生ですが、真核藻類・地衣類・シダ植物・裸子植物などと共生する種もあります。シアノバクテリアは好気生物でしかも光合成は酸素発生型なので、多くの窒素固定シアノバクテリアでは一部の細胞を、光化学系Ⅱを欠いた窒素固定細胞=ヘテロシスト(heterocysts 異型細胞))に分化させることで光合成系と窒素固定系を空間的に分離し、窒素固定と光合成の起こる時間を分離することで酸素感受性の高いニトロゲナーゼを酸素から保護しています。アナベナ(Anabaena)がその代表例です。しかし、嫌気・微嫌気条件でのみ窒素固定活性を発現するレプトリンビア・ボリアナなどの種も存在します。
ソテツ科の植物は9属90種が知られ、これまでにその約1/3に藍藻類が侵入した根粒の着生が報告されています。ソテツの根粒は地表の近くに形成され、多年生で叉状分岐をしており、10cmもの大きさになるものもあります。最近では、ソテツの根粒は、イヌマキの根粒と同様、微生物と関係のない本来の性質であって、藍藻類などの内生菌は2次的に侵入したものと推定されています。
窒素固定メタン菌
深海熱水環境には窒素固定能をもつ好熱性メタン菌が棲息しています。35億年前の深海熱水性の石英脈に保存された窒素分子と(最古のメタン菌由来と考えられている)有機物の窒素同位体組成の関係を調べたところ、当時の深海熱水環境に生息したメタン菌が窒素固定して増殖していた可能性が高いと考えられています。窒素固定遺伝子の大規模な伝播は地球初期の深海熱水環境で起き、生命の共通祖先もしくはメタン菌(当時の深海熱水環境に生息していた)から光合成細菌の祖先に伝播したと考えられています(2014西澤学)。
フランキア菌
フランキアは放線菌門に属するグラム陽性細菌です。フランキアは1886年Brunchorstにより非マメ科植物の根粒中に見出され、その名は彼の師であるスイスの微生物学者A. B. Frankに由来します。フランキアは、多細胞性の菌糸、ベシクル、胞子の3つのタイプの細胞に分化します。通常は、一般的な放線菌と同様に菌糸として生育します。培地中の窒素源が欠乏すると、ベシクルと呼ばれる球状の細胞を分化させ、そこで窒素固定反応を行います。フランキアはベシクルを形成することにより自分自身で酸素防御を行うため、このような好気状態でも窒素固定を行えます。
フランキアが共生する植物は世界で8科14属、総計158種あります。日本ではハンノキ属16、グミ属13、ヤマモモ属3、ドクウツギ属1の計33種について根粒の形成が報告されています。オランダのグルチノザハンノキを主体とした森林では、毎年60~130kg/haの窒素の蓄積が見られます。アメリカのカリホルニア湖では、湖畔に面してハンノキ林が密生していて、湖畔周囲の土壌及び湖水の水が富栄養化し、プランクトンが旺盛に発育しています。ヤマモモは、瀬戸内の石英粗面岩地帯における粘土質土壌の改良に用いられています。マツと混植後12年間に、毎年80kg/haの窒素増加がみられています。
窒素固定エンドファイト(Diazotrophic endophytes)
サトウキビやサツマイモの内部にはエンドファイト細菌が共生しています。エンド(endo)は体内、ファイト(phyte)は植物の意味です。野生イネから分離されたHerbaspirillumはイネの細胞間隙に生息して、窒素固定をします。サツマイモ体内には、窒素固定活性を持つBradyrhizobium属、Pseudomonas属、Paenibacillus属のエンドファイトが生息しています。これらの菌は、土壌にアミノ酸などの有機体窒素があるときに植物体内に入り込みます。化学肥料では入りません。これらの窒素固定エンドファイトは宿主特異性が低く、広範な作物の窒素栄養の改善に利用できます。窒素固定エンドファイトはススキにもあります。
大腸菌は20分以内に一回分裂すると言われています。ヒトの皮膚や肝臓の細胞が入れ替わるのには1ヶ月かかります。大腸菌の方が圧倒的に活動的なのです。
20分に一回分裂すると24時間で72回分裂します。10^X倍に増殖するときの指数Xは
すなわち理論上10^21個以上に増殖します。実際は栄養がなくなって増殖は止まります。仮に栄養が供給され続けると、大腸菌の重量は700fg(=7×10^-13g/個)ですから、1個の細菌は24時間後には、
700トンという途方もない重量になります。細菌には環境を変える力が備わっています。
菌体には、カビと細菌を平均して炭素100gに対し、窒素15g、リン11.6g、カリウム9.8g、カルシウム1.4gが含まれています。つまり菌体にはN:P:K=3:2:2の割合で栄養素が含まれています。菌体のC/N比は6.7です。
糸状菌は炭素100gに対して、窒素5g(4.5g~7.5g)程度含んでいます。糸状菌の場合、
となります。堆肥作製時のC/N比を20とするのはそのためです。堆肥はC/N比10程度まで分解すると、林の匂いがなくなって施用可能になります。ちなみに好気性菌では0.75g~1.5gの窒素を含んでいます。細菌の場合
となります。細菌に必要なのは炭素であることが分かります。
畑10a当たり、700kgの菌体がいると考えられています。乾燥菌体量は、約100kg(=700kg×14.3%)になります。内訳は炭素70kg、窒素11kg、リン8kg、カリ7kg、カルシウム1kgとなります。10aの施肥量は、窒素10kg、P2O5が10kg、K2Oが10kgですから、リンは4.4kg、カリウムが8.3kgとなります。つまり、乾燥菌体のミネラルと施肥量はほぼ等しいといいうことになります。土壌中の菌体が死んで肥料として供給されれば、無肥料でも植物は育つことになります。
好気性細菌は摂取エネルギの5~10%、嫌気性細菌は摂取エネルギの2~5%しか菌体合成に使いません。つまり細菌は摂取エネルギの95%以上を生命維持活動に使ってしまいます(西尾道徳1989年)。細菌を増やすには土壌に大量の炭素が必要なのです。施肥した有機体窒素は、細菌に取り込まれずに余り、硝酸態窒素に変化し、植物に吸収されます。
一方、真核生物であるカビは摂取エネルギの30~50%を菌体合成に使っています。糸状菌などは摂取エネルギの半分を菌糸の伸長に費やしています。糸状菌は細菌よりN、P、Kなどのミネラルが1桁多く必要になります。雑草堆肥などを作るためには、米ぬかなどの窒素分を入れます。糸状菌は湿った土壌を好むので、堆肥が乾かないように時々水を追加して、均一になるように切り返します。
呼吸活性の値の例を挙げると、静止時では
です。正確な単位は[μlO2/mg(乾物)/hour]です。マウスの静止時の呼吸活性がヒトより高いのは、マウスは体が小さいために放熱が大きいからでしょう。活動時は上昇し
となります。蝶は飛翔するのに静止時の166倍のエネルギを使っています。
細菌の呼吸活性はどうでしょうか?
大腸菌の呼吸活性はヒトより100倍以上大きいことが分かります。窒素固定菌に至っては1000倍以上です。細菌を増やすには土壌に大量の有機物が必要になりそうです。
体重65kgの18才男子の基礎代謝量は
です。1分当たりに換算すると、1日は1440分(=24×60)なので
となります。安静時の酸素消費量は約230[ml/min]です。1kg当たりの酸素消費量は
となります。METSはスポ-ツ生理学で用いられている運動強度の単位です。つまりエネルギ消費量と酸素消費量の比は
です。つまり酸素消費量1[ml]当たり5[cal]のエネルギを生み出しています。
従って、エネルギ消費量Xと酸素消費量Yの間には
の関係が成り立っています。1 METSの消費エネルギは
です。
呼吸活性とは、1時間に重量1mg当たりの酸素消費量(μl:マイクロ・リットル)のことです。呼吸活性が大きいほど生命活動が活発だと言えます。
ヒトの安静時の酸素消費量は約230[ml/min]です。1kg当たりの酸素消費量は
となります。
歩行時には5倍、走行時には10倍の酸素消費量になるので、
となります。
ヒトの細胞の呼吸活性は細胞によって異なり
となっています(1980年柳田)。全体の酸素消費量0.21に比べて、上記の器官は活発に活動していることが分かります。