参考文献:Joao C. Setubal, Virginia
Bioinformatics Institute, JOURNAL OF BACTERIOLOGY, July 2009, p. 4534–4545,’Genome
Sequence of Azotobacter vinelandii, an Obligate Aerobe Specialized To Support
Diverse Anaerobic Metabolic Processes’
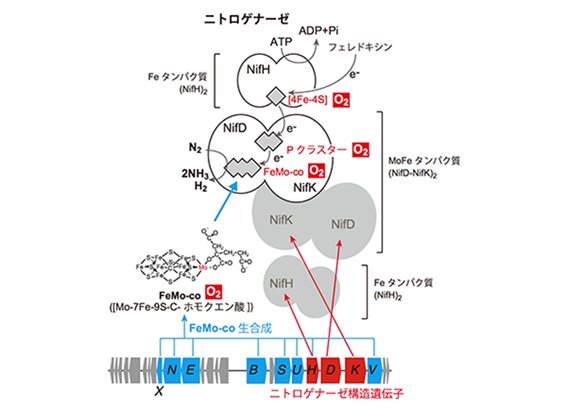
Fe タンパク質にある[4Fe-4S]クラスタは4つの鉄と4つの硫黄がキュバン(cubane)状(=立方体状)に集合したFe-Sクラスタです。MoFe タンパク質は、[8Fe-7S]構造のPクラスタとFeMo-coと呼ばれる[Mo-7Fe-9S-C-ホモクエン酸]の有機金属クラスタを含み、NifDとNifKの2つのサブユニットによるα2β2というヘテロ4量体構造をしています。[4Fe-4S]クラスタから送られてきた電子は、Pクラスタを経由してFeMo-co(フェモコ)に伝達され、FeMo-co上に結合した窒素分子を還元します。PクラスタからFeMo-coに電子を伝達すると、
P cluster(還元型)+FeMo-co(酸化型)→ P cluster(酸化型)+FeMo-co(還元型)
窒素固定菌はニトロゲナーゼ(Nitrogenase)という酵素を使って、窒素分子をアンモニアへと変換します。炭化水素のC-H間の結合エネルギは約100 kcal/mol程度であるのに対して、N≡Nの三重結合のエネルギは225
kcal/molと高いので、これを還元的に開裂して2分子のアンモニアに変換することは大変なことです。Hoffman教授はニトロゲナーゼを“Everest of enzyme”と呼んだそうです(2009年)。ニトロゲナーゼは、モリブデンMoを含むMo型、バナジウムVを含むV型それに鉄Feのみを含むFe型の三種類あります。特に、土壌細菌のアゾトバクタ・ヴィネランディ(vinelandii)のMo型ニトロゲナーゼがモデル細菌となっています。ニトロゲナーゼは以下のような反応を触媒します。ここでPiはリン酸を表します。
フランキアは放線菌門に属するグラム陽性細菌です。フランキアは1886年Brunchorstにより非マメ科植物の根粒中に見出され、その名は彼の師であるスイスの微生物学者A. B. Frankに由来します。フランキアは、多細胞性の菌糸、ベシクル、胞子の3つのタイプの細胞に分化します。通常は、一般的な放線菌と同様に菌糸として生育します。培地中の窒素源が欠乏すると、ベシクルと呼ばれる球状の細胞を分化させ、そこで窒素固定反応を行います。フランキアはベシクルを形成することにより自分自身で酸素防御を行うため、このような好気状態でも窒素固定を行えます。