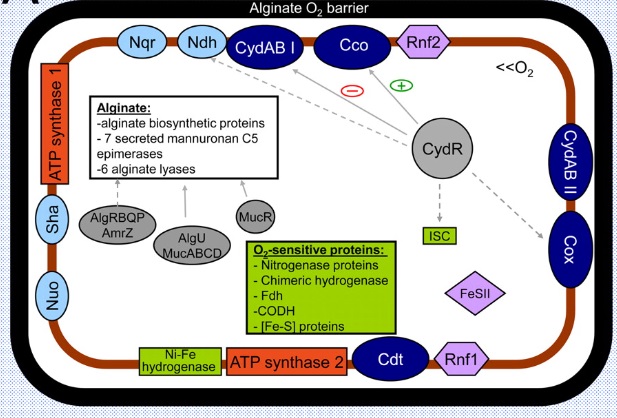
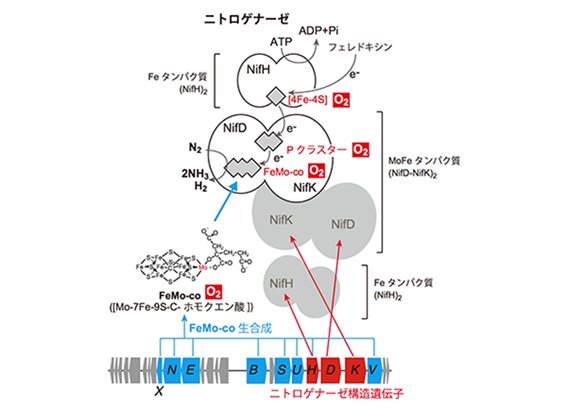
窒素固定菌というとマメ科の根粒菌(Rhizobium)が有名ですが、他にも単体で窒素を固定するアゾトバクタ(Azotobacter)、クロストリジウム(Clostridium)、藍藻(diazotrophic cyanobacteria)がいます。またフランキア(Frankia)などの放線菌(Actinomycetales)は多くの樹木と共生し、空気中の窒素をアンモニアに変えて摂取しています。土壌の中のアゾトバクタが十分いれば、作物の幼苗は、窒素肥料を殆ど与えなくても、成長します。窒素肥料を与えてしまうと、アゾトバクタと作物との緩い共生関係は無くなってしまいます。牛糞堆肥などを使う有機農法から無肥料栽培に転換するには、土壌中の窒素成分を減らして、微生物の転換を図らなければなりません。
マメ科の根粒菌
マメ科植物の多くは根に粒状の根粒を形成し、そのなかに根粒菌が共生しています。根粒菌は植物から糖をもらい、植物に空気中の窒素を分解して得た窒素化合物やホルモンを供給しています。マメ科植物は世界各地に分布しており、現在450属、13,000種ほどが知られています。調査された2,000種のうち、根粒を形成しない種類が約10%ありました。
アルファルファ、クロ-バ、エンドウ豆、インゲン豆、ル-ビン、大豆、カウピ-(落花生)、ミヤコグサ、ダレヤ、イガマメ、ニセアカシア、イタチハギ、タチレンゲソウ、ムレスズメなど約20種類のマメ科の植物は、交互に根粒菌を交換しても根粒が形成されます。根粒菌は、土壌中では鞭毛のある小型の球菌ですが、共生する時は桿状大型化し、不規則な形態のバクテロイドになります。根粒組織中にはレグヘモグロビンという赤色の色素がみられます。根粒の寿命は多くは1年以内であって、結実するころから根粒の内容物は寄主植物に吸収され、根粒内の根粒菌は土中に放出されます。
根粒の形成には、好気的条件が必要です。また窒素肥料が多いと根粒が形成されません。リン酸は根粒形成に不可欠で、根粒形成を促進させます。微量元素の硼素(B)は根粒とバクテロイドの形成に、モリブデン(Mo)は窒素固定に必須の元素です。マメ科作物の種子に根粒菌を接種して根粒を形成させる人工接種の手法が開発されています。人工接種すると、無効菌が先に根に侵入する前に根粒が形成されるので、作物の収穫量と品質が向上するようです。
アゾトバクタ
アゾトとはイタリア語で窒素の意味です。クロオコッカムやビネランジは、シュ-ドモナス科アゾトバクタ属のグラム陰性の好気性細菌です。アゾトバクタ属の細菌は単体で自分のために窒素を空気中から取り入れ固定します。アゾトバクタは1gの炭水化物を消費して5~20mgの窒素を生産します。これは根粒菌の10%程度です。
アゾトバクタは、土壌中に広く分布し、中性付近で窒素固定を行うため、酸性土壌にはあまりいません。植物は根から糖を出し、アゾトバクタはアンモニウムを出して、お互いに緩い共生関係を保っています。特にアゾトバクタは光合成細菌と共生します。アゾトバクタは脂肪酸を提供し、光合成細菌は糖を提供します。そのため光合成細菌を施肥すると窒素固定量が増えます。アゾトバクタは大量の酸素を消費するので、環境が嫌気的になりがちですが、光合成細菌は酸素がなくてもATPを生産できるので、アゾトバクタと共生できるのです。
窒素固定細菌に関しては、アゾトバクター属以外に4属が知られています。
- アグロモナス属Agromonas 酸素分圧の低いところで窒素固定する。
- アゾモナス属Azomonas 低いpH(4.6~4.8)域で窒素固定する。
- ベイゼリンキア属Beijerinckia 37℃で生育、熱帯地方で窒素固定する。
- デルキシア属Derxia メタンを同化でき、熱帯地方に分布し窒素固定する。
クロストリジウム
クロストリジウムは3~4μmサイズのグラム陽性の嫌気性細菌です。繊毛があり運動します。耐酸性があり、あらゆる土壌に分布しています。但し窒素固定力はアゾトバクタより弱いです。クロストリジウム属細菌は、SODやカタラーゼなどの活性酸素を無毒化する酵素を持たないため、酸素がある通常の環境下では不活化します。酸素存在下では、耐久性の高い芽胞を作って休眠することで、死滅を免れます。ボツリヌス菌や破傷風菌やウエルシュ菌はクロストリジウム属の細菌です。サ-モセラム菌は好熱性で酸素なしにセルロ-スを分解できるため、エタノール生産に利用されています。クロストリジウム属菌はガン細胞を選択的に攻撃することが知られており、その応用が研究されています。
窒素固定シアノバクテリア
単細胞・糸状体種のシアノバクテリアの半数は窒素固定の能力を持ちます。大部分は単生ですが、真核藻類・地衣類・シダ植物・裸子植物などと共生する種もあります。シアノバクテリアは好気生物でしかも光合成は酸素発生型なので、多くの窒素固定シアノバクテリアでは一部の細胞を、光化学系Ⅱを欠いた窒素固定細胞=ヘテロシスト(heterocysts 異型細胞))に分化させることで光合成系と窒素固定系を空間的に分離し、窒素固定と光合成の起こる時間を分離することで酸素感受性の高いニトロゲナーゼを酸素から保護しています。アナベナ(Anabaena)がその代表例です。しかし、嫌気・微嫌気条件でのみ窒素固定活性を発現するレプトリンビア・ボリアナなどの種も存在します。
ソテツ科の植物は9属90種が知られ、これまでにその約1/3に藍藻類が侵入した根粒の着生が報告されています。ソテツの根粒は地表の近くに形成され、多年生で叉状分岐をしており、10cmもの大きさになるものもあります。最近では、ソテツの根粒は、イヌマキの根粒と同様、微生物と関係のない本来の性質であって、藍藻類などの内生菌は2次的に侵入したものと推定されています。
窒素固定メタン菌
深海熱水環境には窒素固定能をもつ好熱性メタン菌が棲息しています。35億年前の深海熱水性の石英脈に保存された窒素分子と(最古のメタン菌由来と考えられている)有機物の窒素同位体組成の関係を調べたところ、当時の深海熱水環境に生息したメタン菌が窒素固定して増殖していた可能性が高いと考えられています。窒素固定遺伝子の大規模な伝播は地球初期の深海熱水環境で起き、生命の共通祖先もしくはメタン菌(当時の深海熱水環境に生息していた)から光合成細菌の祖先に伝播したと考えられています(2014西澤学)。
フランキア菌
フランキアは放線菌門に属するグラム陽性細菌です。フランキアは1886年Brunchorstにより非マメ科植物の根粒中に見出され、その名は彼の師であるスイスの微生物学者A. B. Frankに由来します。フランキアは、多細胞性の菌糸、ベシクル、胞子の3つのタイプの細胞に分化します。通常は、一般的な放線菌と同様に菌糸として生育します。培地中の窒素源が欠乏すると、ベシクルと呼ばれる球状の細胞を分化させ、そこで窒素固定反応を行います。フランキアはベシクルを形成することにより自分自身で酸素防御を行うため、このような好気状態でも窒素固定を行えます。
フランキアが共生する植物は世界で8科14属、総計158種あります。日本ではハンノキ属16、グミ属13、ヤマモモ属3、ドクウツギ属1の計33種について根粒の形成が報告されています。オランダのグルチノザハンノキを主体とした森林では、毎年60~130kg/haの窒素の蓄積が見られます。アメリカのカリホルニア湖では、湖畔に面してハンノキ林が密生していて、湖畔周囲の土壌及び湖水の水が富栄養化し、プランクトンが旺盛に発育しています。ヤマモモは、瀬戸内の石英粗面岩地帯における粘土質土壌の改良に用いられています。マツと混植後12年間に、毎年80kg/haの窒素増加がみられています。
窒素固定エンドファイト(Diazotrophic endophytes)
サトウキビやサツマイモの内部にはエンドファイト細菌が共生しています。エンド(endo)は体内、ファイト(phyte)は植物の意味です。野生イネから分離されたHerbaspirillumはイネの細胞間隙に生息して、窒素固定をします。サツマイモ体内には、窒素固定活性を持つBradyrhizobium属、Pseudomonas属、Paenibacillus属のエンドファイトが生息しています。これらの菌は、土壌にアミノ酸などの有機体窒素があるときに植物体内に入り込みます。化学肥料では入りません。これらの窒素固定エンドファイトは宿主特異性が低く、広範な作物の窒素栄養の改善に利用できます。窒素固定エンドファイトはススキにもあります。