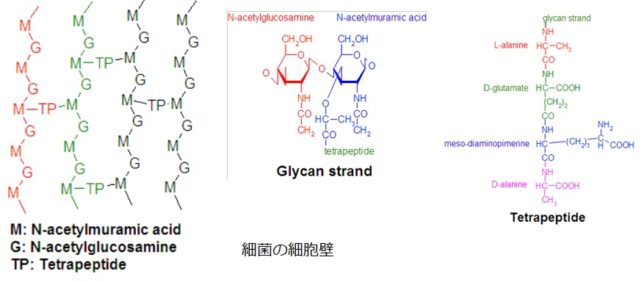
植物の細胞壁は、セルロ-ス、ペクチン、ヘミセルロ-スなどの繊維が架橋性多糖やタンパク質で絡み合い、リグニンで固化された構造をしています。強固な細胞壁のおかげで植物は起立しています。そのためよくセルロ-スは鉄筋、ペクチンはコンクリ-トに例えられますが、細胞壁は生きた組織です。なぜなら細胞壁の中には様々な酵素タンパク質が含まれており、細胞の中で起こる生命活動に深く関わっているからです。
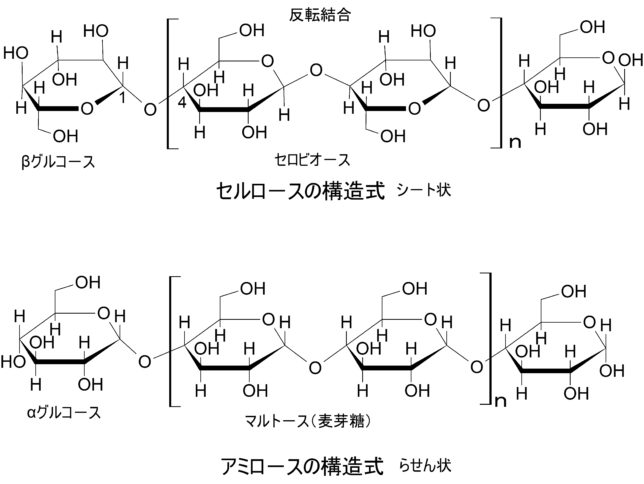
セルロースやペクチンは糖の鎖です。セルロースとはD-グルコースという糖がβ-1,4-結合で長く結合した高分子です。グルコ-スにはα型とβ型があります。各々の OH 基のグルコ-ス環に対する上下関係で区別します。C1とC4に結合したOH基がグルコ-ス環に対して同じ側に突き出しているのがα型です。αグルコ-スが鎖状に結合すると水素結合のせいでラセン型のアミロ-ス(デンプン)になります。アミロ-スはマルト-ス(麦芽糖)が重合した構造をしています。βグルコ-スが鎖状に結合するとシート型のセルロ-スになります。セルロ-スはラセン型でないので、ヨウ素デンプン反応を示しません。セルロ-スはセルビオ-スが重合した構造をしています。セルビオ-スは2つのβグルコ-スの片方が裏返されて結合しています。面白いですね。
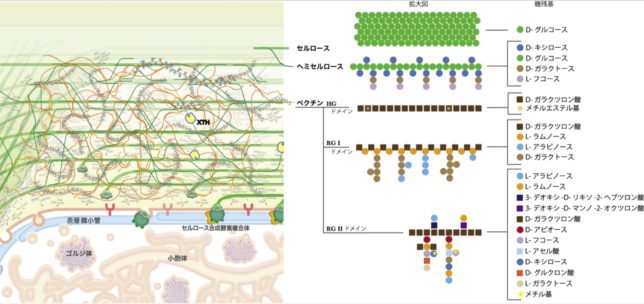
セルロースの構成する細胞壁繊維は微繊維の集合体から成ります。微繊維は結晶性ミセルが数個集まった構造で直径は30nmです。結晶性ミセルはセルロース分子40本が水素結合で束ねられた構造体(直径5nm)です。微繊維間隙幅は10nmで、この空隙にはキシログリカンなどのヘミセルロースが満たされており、微繊維間の構造的強度を高めています。
図1に植物の一次細胞壁の構造モデルを示します。色とりどりの●と■は多糖類を構成する糖残基を示しています。一次細胞壁は、結晶性のセルロース微繊維がヘミセルロースにより架橋された網状構造が骨格となり、その隙間を巨大分子であるペクチンが埋める構造モデルが広く受け入れられています。
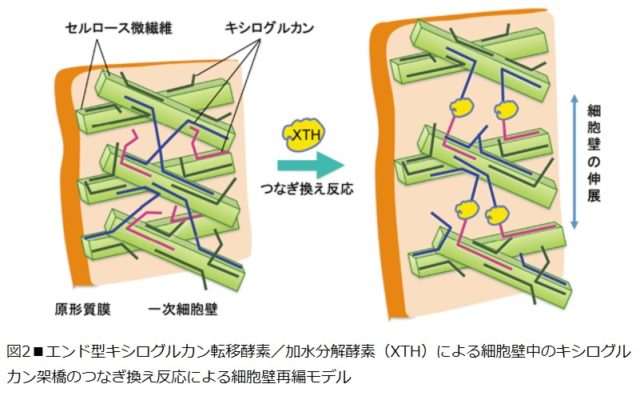
セルロース微繊維は、細胞膜上のセルロース合成酵素により合成されます。合成酵素は細胞内の表層微小管に沿って動くため、微繊維の向きは表層微小管により決定されます。ヘミセルロースやペクチンは、ゴルジ体内で多数の膜貫通型の糖転移酵素の働きで合成された後、膜交通を介して細胞壁中に分泌されます。細胞壁中では、それぞれXTH(=Xyloglucan endoTransglucosylase/Hydrolase)やPME(=Pectin Methyl Esterase)などによる修飾を受けながら、細胞壁の高次構造に組み込まれます。
図2にエンド型キシログルカン転移酵素・加水分解酵素(XTH)による細胞壁中のキシログルカン架橋のつなぎ換え反応による細胞壁再編モデルを示します。つなぎ換え反応の際に切断されるキシログルカン鎖(供与鎖)を青色,つながれるキシログルカン鎖(受容鎖)を赤色で表示しています。細胞壁が伸展するのは、XTHによるつなぎ換え反応により、一次細胞壁のセルロースとキシログルカン網状構造の再編が起こるからです。
ペクチンはポリ・ガラクツロン酸の重合体です。ガラクツロン酸(galacturonic acid)はガラクトースが酸化されたウロン酸です。ガラクトースはC2とC5のOH基が同じ方向を向いている糖です。C5にCH2OHがついているのがガラクトースで、C5にCOOHがついているのがウロン酸です。ちょっとした違いですが、糖鎖の化学は立体異性体の区別が難しいです。
図3にペクチン・メチル・エステラーゼ(PME)による脱メチル化を介したホモガラクツロナン(=HG)のカルシウム架橋の形成モデルを示します。
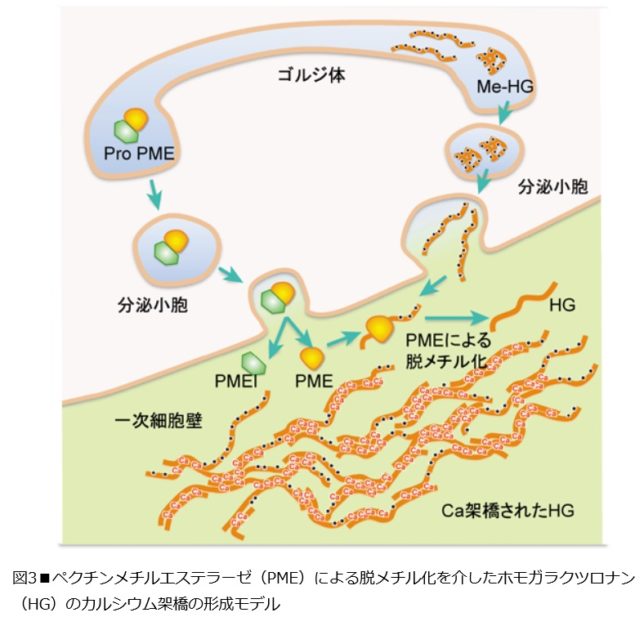
ペクチンの全ドメイン内で最も大きな領域を占めるHGドメインは、ガラクツロン酸がα-(1→4)-グリコシド結合した直鎖状の多糖として細胞内のゴルジ体で合成されると同時に、ガラクツロン酸残基中のカルボキシル基がメチルエステル化され、電荷をもたない状態で細胞壁中へ分泌されます。一方、PMEは不活性な前駆体(Pro-PME)としてゴルジ体を経て分泌される過程で不活性化ドメイン(PMEI)が切り離され、活性型PMEとなります。 メチルエステル化されたMe-HGは細胞壁中に分泌された後、PME酵素により脱メチル化されます。脱メチル化されることで、HGはCaイオンを介して高度な分子間架橋を形成してゲル化し、植物の細胞に強度と柔軟性を与えます。ペクチンのRGIIドメインは側鎖中のアピオース残基のジオール基間で、ホウ素(B)を介して分子間架橋を形成します。つまりCaやBが植物の必須元素である理由のひとつは、細胞壁を形成するのに不可欠な元素だからです。
ヘミセルロースはキシランやマンナンのほか、グルコマンナンやグルクロノ・キシランなどのような複合多糖もあります。ヘミセルロースは結晶性セルロース微繊維同士の凝集を防ぎ、細胞壁の伸展性を高めています。また化学的に安定な結晶性セルロース微繊維の表面を多様な分子種からなるヘミセルロースで覆うことで、化学反応を伴う細胞壁の形質変化を可能にしています。
多くの被子植物の細胞壁はタイプIと呼ばれ、セルロース、ペクチン、キシログルカンが多く含まれています。イネ目の細胞壁はタイプIIと呼ばれ、セルロース、キシラン、βグルカンが多く、ペクチンやキシログルカンが少ないです。タイプIIではタンパク質含量が低く、代わりにフェノール酸の架橋がその役割を果たしています。
参考文献 横山隆亮他、化学と生物53巻No.2 P107-114 (2015)「植物細胞壁: 高次構造の構築と再編」、東北大学大学院生命科学研究科