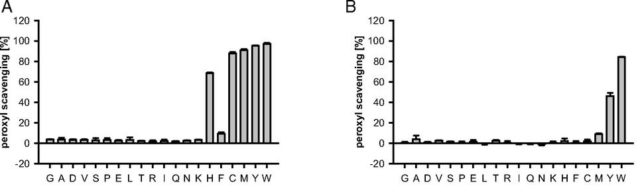
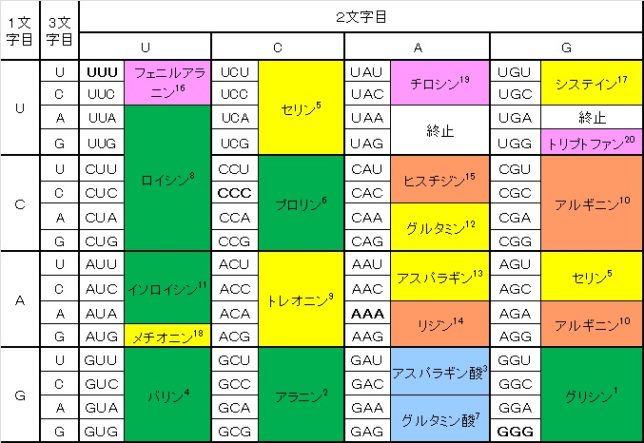
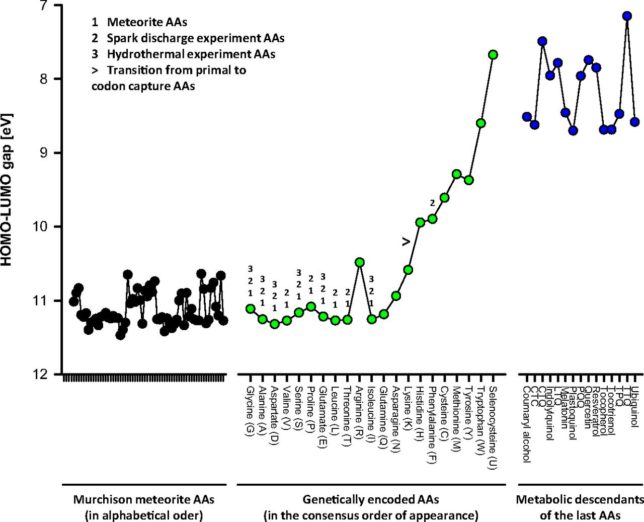
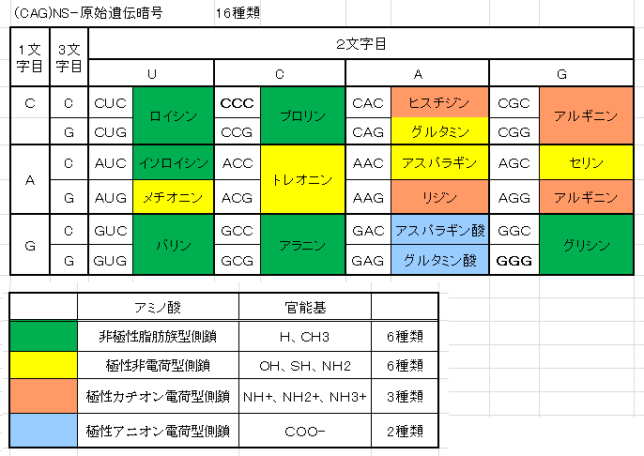
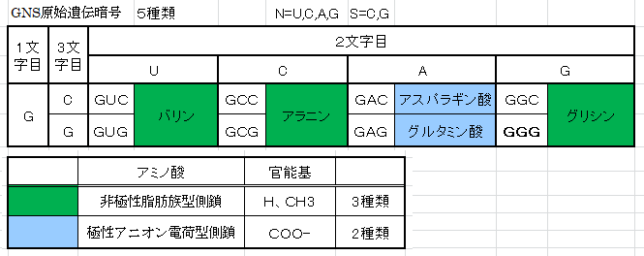
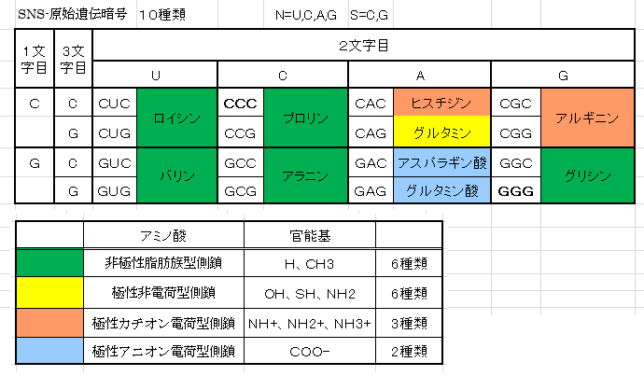
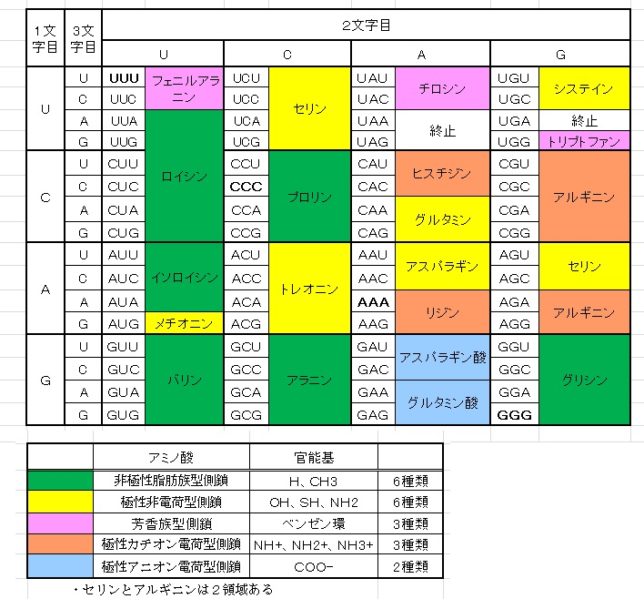
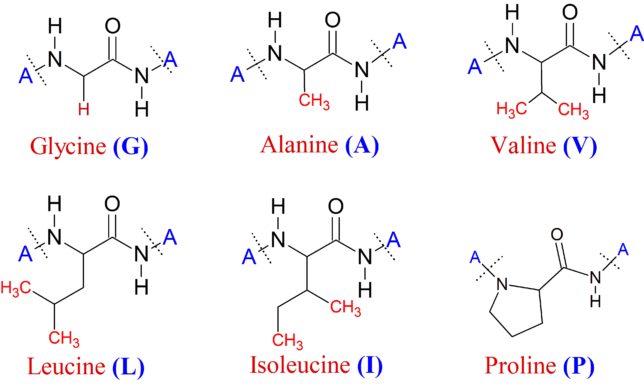
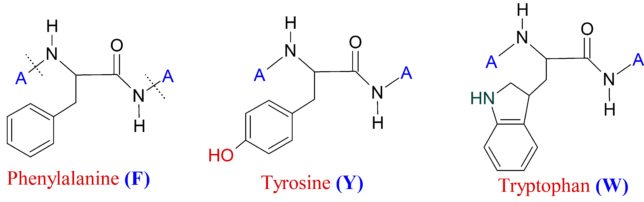
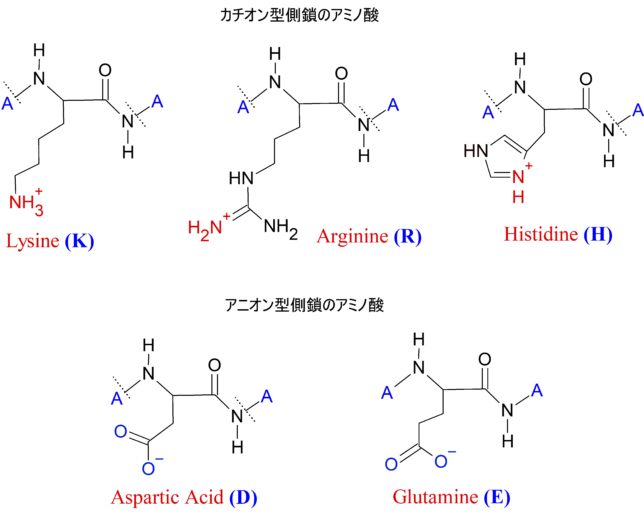
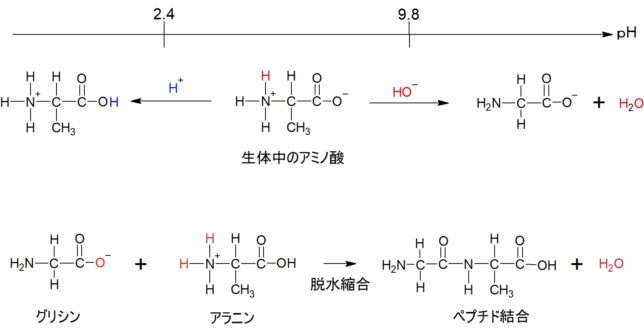
牛乳には100cc当たり100mgのCaが含まれています。しかしCaの吸収率は低く、牛乳で50%程度、ほかの食品では20~30%程度しかありません。年齢とともに吸収率が悪くなるので、中高年の場合にはCa摂取量の目標を1000mg/日にして、できるだけ和食を食べるようにしましょう。一時期、「牛乳を摂ると骨が弱くなる」と言われたことがありましたが、科学的には根拠がないようです。
Caを吸収するには、ビタミンDと紫外線(日光浴)、Mg(マグネシウム)が必要です。ビタミンKも必要ですが、これは体内で再生産されるので欠乏しません。ビタミンDには、紫外線によって皮膚でコレステロール前駆体から生合成されるビタミンD3 (コレカルシフェロール)と、食物から摂取されるビタミンD2(エルゴカルシフェロール)の2種類がありますが、両方の代謝経路と作用は殆ど同じです。 ビタミンDは下部小腸から吸収され、蛋白質と結合して血中を運ばれ、まず肝臓において25位が水酸化されて、25(OH)Dへと変換されます。さらに、腎臓近位尿細管の1α水酸化酵素によって1α位が水酸化され、最終的な活性型ホルモンである1,25(OH)2Dが生成されます。活性型ビタミンDは、Caを骨に届けて固定するまでを手助けします。小魚はCaとビタミンDを含みますが、牛乳にはビタミンDは含まれていません。
ビタミンD2 はキノコなどの植物性食品に含まれ、ビタミンD3 は魚類や鶏卵などの動物性食品に含まれます。中華料理に使うキクラゲ(キノコ)1個(1g)にはビタミンD2が4.3μgもはいっており、2個食べるだけで、1日分の摂取量を賄えます。しらすは大さじ2杯(6μg)で1日分の摂取量が得られます。
Caを吸収するには、Mgとのバランスも重要だと言われています。理想の割合は、Ca:Mg=2:1だそうです。Mgを多く含むのは、大豆製品、海藻、緑黄色野菜、ナッツ類などです。Mgの必要量は1日あたり300mgで、Caの半分程度です。

塩分の摂りすぎ
忘れがちなのが塩分です。塩分(NaCl)を多くとるとCaの吸収が阻害されると言われています。これはCaCl2が腸で析出して排出されるのではなく、Naの増加によって腎臓でのCaの再吸収が阻害されるということです。もう少し詳しく説明します。
腎臓の糸球体からは遊離Ca2+イオン(濃度5mg/dl)が1日5,000mgも濾過されています。濾過されたCaの殆どは再吸収されて、1日100mg強のCaのみが尿中に排泄されます。濾過されたCaの55%は近位尿細管で、25%がヘンレ係蹄、15%が遠位尿細管で、さらに5%が集合管で再吸収されます。つまり糸球体から濾過されたCaの95%以上が腎尿細管で再吸収されます。このうち近位尿細管でのCa再吸収は、Na輸送に伴う受動的なものであり、ヘンレ上行脚でのCa再吸収もClイオンの輸送に伴う電位差に関連した二次的なものです。
これに対し遠位尿細管でのCa再吸収はNa再吸収や電位差に依存せず、PTHの作用により調節を受けます。つまりNaClの摂取量が増加すると、Ca再吸収の80%を占める尿細管(近位尿細管とヘンレ係蹄)でのCa再吸収が阻害されるというわけです。ちなみに活性ビタミンDは遠位尿細管でのCa再吸収を自ら高めるとともに、PTHのカルシウム再吸収を増強します。
私たちが食塩を摂取するのは、野菜自体には塩分が乏しいからなのでしょう。野菜の塩漬は保存をよくするばかりでなく、うまみ味や塩分を補うよい方法です。ヒトの血液には0.85%の塩分が含まれています。料理の味付けはその濃度に近くするといいようです。食塩は1日に7~8gでよいのですが、日本人は平均して2~3g過剰に食塩を摂取しています。塩分の摂りすぎ自体も血管や腎臓に負担をかけることになります。
ごはんには塩は入っていませんが、パンやチ-ズやハムには塩分が入っています。スナック菓子の塩分も多いです。外食が多い人は、塩分の摂りすぎに注意が必要です。運動をする人は汗で塩分が除去されますが、運動量の少ない人は、ラーメンやうどんの汁を残すようにして、塩分の摂り過ぎを避けるといいでしょう。
エストロゲンとCa
女性は妊娠や授乳期などCaを大量に必要とする時期があります。また閉経期を迎えると、卵巣から分泌される女性ホルモンのエストロゲンが急激に減少します。エストロゲンは、骨からカルシウムが溶け出すのを防ぐ役割を担っているので、エストロゲン量が低下すると、骨がもろくなりやすいのです。また血中Caを血管壁が吸収し、血管が固くなるので、閉経後の女性は血圧が上昇しやすくなります。エストロゲンの減少と塩分の摂りすぎが重なると、腎臓でのカルシウムの再吸収量が減って、尿と一緒にCaが排出されてしまいます。
48歳過ぎからは、食事と運動を習慣づけるといいでしょう。子どもが独立する年齢なので、寂しくなります。子犬を飼って散歩につれていくのはいいことなのかもしれません。
P(リン)の過剰摂取について
Caの推奨摂取量は800mg/日程度で、上限量は2500mg/日程度でした。Pの推奨摂取量は1000mg/日程度で、成人のPの耐容上限量は3000mg/日となっています。この量にできるだけ近づかない注意が必要です。骨の構成物質であるハイドロキシアパタイトCa10(PO4)6(OH)2には、P:Ca=6:10の割合で含まれています。つまり骨にはPより多くのCaが必要です。
リン酸塩とpH調整剤の表示があるソーセージのP摂取量は180±70mg/100g(2015年)です。1食200gのソ-セ-ジには約400mgのPが含まれていると考えていいでしょう。加工食品には上限値にならない程度にリン酸塩が添加されています。しかしリン酸はあまりに多くの加工食品に使われているので過剰摂取が心配されています。
法律では加工食品に用途の記載義務はありますが、添加物名の記載義務はありません。しかし加工食品の材料欄に、結着剤、乳化剤、酸味料、pH調整剤といった用途が記載されていればリン酸塩が含まれています。ハムやソーセージのプリッとした食感はリン酸結着剤によるものです。プロセスチーズの乳化剤は複数のチーズを均一に混ぜるために用いられます。ファミレスのドリンクバーで供給されるコーヒーの抽出液にはリン酸が入っています。お湯で抽出するよりも3倍も多く抽出ができるからです。失われた香りは人工的に添加されています。ラ-メンの麺の食感を作り出すかんすいにもリン酸塩が含まれています。缶詰、佃煮、煮豆、味噌の変色防止やpH調整のためにもリン酸塩が用いられています。添加物の摂り過ぎが体に良くない理由を理解した上で、上手に添加物と付き合いましょう。