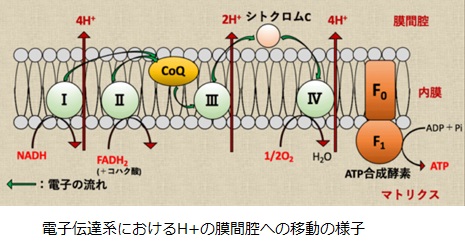
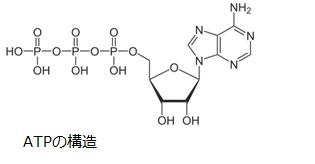
結局NADHを電子伝達系で用いる場合には、合計10H+がマトリクスから膜間腔へ輸送されます。FADH2の場合は合計6H+が膜間腔へ輸送されます。膜間腔のH+濃度と電位が高くなっているので、ミトコンドリア内膜を挟んだプロトン駆動力を利用してATP合成酵素がATPを合成します。
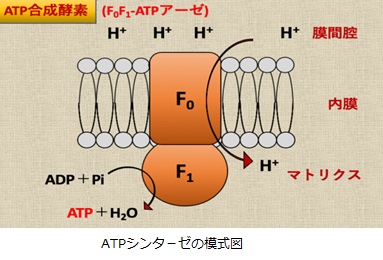
ATP合成酵素はF0サブユニットとF1サブユニットによる分子モ-タとして機能します。F0はミトコンドリア内膜に埋まっていて、F1はミトコンドリア・マトリクスに突き出た形で存在しています。F0モーターがH+の濃度勾配によるエネルギを使ってF1モーターを回すことによって、ATPを産生しています。3分子のH+がマトリクスへと輸送されるごとにATP1分子が合成されます。
しかし、実際にマトリクスにおいてATPを合成するためには、ATP合成の材料となるADPやリン酸Piをマトリクス内に取り込む必要があります。また、合成されたATPの大部分は細胞質で利用されるため、マトリクスから細胞質へと輸送される必要があります。
内膜を隔てたATP、ADP、Piの輸送
ミトコンドリア内膜を隔てたATP、ADP、 Piの輸送はアデニン・ヌクレオチド・トランスロカーゼとリン酸輸送体という2つの膜タンパク質によって行われています。アデニン・ヌクレオチド・トランスロカーゼは、ATP-ADP交換タンパク質のことで、細胞質のADP3-をミトコンドリア内へ、ミトコンドリア内のATP4-を細胞質へと対向輸送しています。この対向輸送では、プロトン勾配の電荷の差が用いられています。
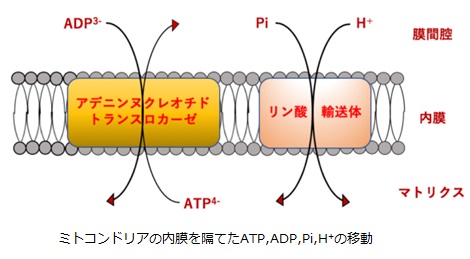 リン酸輸送体は、細胞質のPiをミトコンドリア内へと輸送するときにH+も同時にミトコンドリア内へと共輸送します。この共輸送ではH+の濃度差が用いられています。ちなみに、対向輸送とは、膜の内外で異なる物質を相互に逆方向に移動させる輸送のことで、共輸送とは、膜の片側から異なる物質を同方向に移動させる輸送のことをいいます。
リン酸輸送体は、細胞質のPiをミトコンドリア内へと輸送するときにH+も同時にミトコンドリア内へと共輸送します。この共輸送ではH+の濃度差が用いられています。ちなみに、対向輸送とは、膜の内外で異なる物質を相互に逆方向に移動させる輸送のことで、共輸送とは、膜の片側から異なる物質を同方向に移動させる輸送のことをいいます。
ミトコンドリアの外にATPを輸送し、マトリクスにADPを供給するためには、H+1個分のプロトン駆動力が用いられていました。ATP合成酵素は3H+で1個のATPを産出するので、細胞質内でATPを1分子増やすためには、4個分のH +が膜間腔からマトリクスに流入する必要があります。この数はNADHやFADH2が1分子あたりでどれくらいのATPを産生するかの指標となります。
細胞質内のATPを1分子増やすためには、4個分のH +が膜間腔からマトリクスに流入するということを踏まえると、NADH1分子あたり
・10[H+]/4[H+/ATP]=2.5ATP
FADH21分子あたり
・6[H+]/4[H+/ATP]=1.5ATP
が合成されることになります。
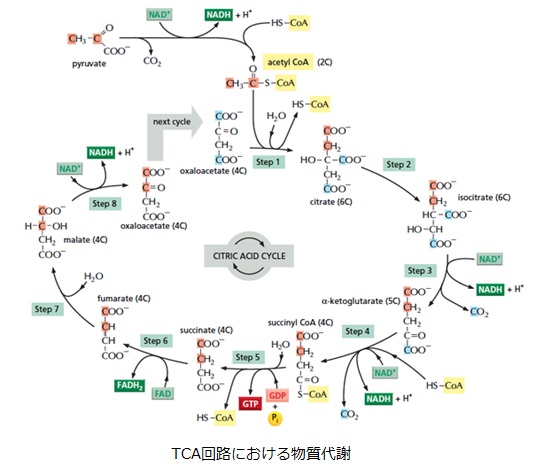
好気呼吸では、1分子のグルコースが「解糖系→ピルビン酸のアセチルCoAへの変換→クエン酸回路」という経路でATPやNADH、FADH2を生成していました。解糖の過程で2分子のATPと2分子のNADH、2ピルビン酸→2アセチルCoAの過程で2分子のNADH、クエン酸回路の過程で2分子のATP(GTP)と6分子のNADHと2分子のFADH2が生成されます。
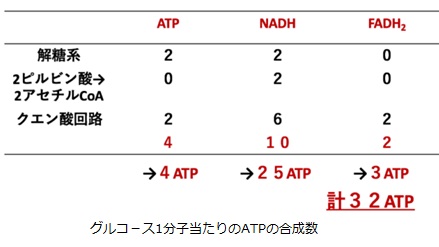
・4ATP+10NADH・2.5[ATP/NADH]+2FADH2・1.5[ATP/FADH2]=4+25+3=32ATP
結局1分子のグルコ-スは、嫌気的代謝では2分子のATPしか生成できませんが、好気的代謝では32分子ものATPを細胞外に生成できることが分かります。
細胞は糖を取り込み、ミトコンドリアで大量のATPを合成できることが分かりました。青森出身の安保先生によると、若い時は解糖系の瞬発力を主に使い、老年期になるとミトコンドリア系の持久力を主に使って活動するので、年齢ともに少食にしていった方が適正体重を保ちやすいそうです。